Educational resources and simple solutions for your research journey


How to Write Review of Related Literature (RRL) in Research

A review of related literature (a.k.a RRL in research) is a comprehensive review of the existing literature pertaining to a specific topic or research question. An effective review provides the reader with an organized analysis and synthesis of the existing knowledge about a subject. With the increasing amount of new information being disseminated every day, conducting a review of related literature is becoming more difficult and the purpose of review of related literature is clearer than ever.
All new knowledge is necessarily based on previously known information, and every new scientific study must be conducted and reported in the context of previous studies. This makes a review of related literature essential for research, and although it may be tedious work at times , most researchers will complete many such reviews of varying depths during their career. So, why exactly is a review of related literature important?
Table of Contents
Why a review of related literature in research is important
Before thinking how to do reviews of related literature , it is necessary to understand its importance. Although the purpose of a review of related literature varies depending on the discipline and how it will be used, its importance is never in question. Here are some ways in which a review can be crucial.
- Identify gaps in the knowledge – This is the primary purpose of a review of related literature (often called RRL in research ). To create new knowledge, you must first determine what knowledge may be missing. This also helps to identify the scope of your study.
- Avoid duplication of research efforts – Not only will a review of related literature indicate gaps in the existing research, but it will also lead you away from duplicating research that has already been done and thus save precious resources.
- Provide an overview of disparate and interdisciplinary research areas – Researchers cannot possibly know everything related to their disciplines. Therefore, it is very helpful to have access to a review of related literature already written and published.
- Highlight researcher’s familiarity with their topic 1 – A strong review of related literature in a study strengthens readers’ confidence in that study and that researcher.

Tips on how to write a review of related literature in research
Given that you will probably need to produce a number of these at some point, here are a few general tips on how to write an effective review of related literature 2 .
- Define your topic, audience, and purpose: You will be spending a lot of time with this review, so choose a topic that is interesting to you. While deciding what to write in a review of related literature , think about who you expect to read the review – researchers in your discipline, other scientists, the general public – and tailor the language to the audience. Also, think about the purpose of your review of related literature .
- Conduct a comprehensive literature search: While writing your review of related literature , emphasize more recent works but don’t forget to include some older publications as well. Cast a wide net, as you may find some interesting and relevant literature in unexpected databases or library corners. Don’t forget to search for recent conference papers.
- Review the identified articles and take notes: It is a good idea to take notes in a way such that individual items in your notes can be moved around when you organize them. For example, index cards are great tools for this. Write each individual idea on a separate card along with the source. The cards can then be easily grouped and organized.
- Determine how to organize your review: A review of related literature should not be merely a listing of descriptions. It should be organized by some criterion, such as chronologically or thematically.
- Be critical and objective: Don’t just report the findings of other studies in your review of related literature . Challenge the methodology, find errors in the analysis, question the conclusions. Use what you find to improve your research. However, do not insert your opinions into the review of related literature. Remain objective and open-minded.
- Structure your review logically: Guide the reader through the information. The structure will depend on the function of the review of related literature. Creating an outline prior to writing the RRL in research is a good way to ensure the presented information flows well.
As you read more extensively in your discipline, you will notice that the review of related literature appears in various forms in different places. For example, when you read an article about an experimental study, you will typically see a literature review or a RRL in research , in the introduction that includes brief descriptions of similar studies. In longer research studies and dissertations, especially in the social sciences, the review of related literature will typically be a separate chapter and include more information on methodologies and theory building. In addition, stand-alone review articles will be published that are extremely useful to researchers.
The review of relevant literature or often abbreviated as, RRL in research , is an important communication tool that can be used in many forms for many purposes. It is a tool that all researchers should befriend.
- University of North Carolina at Chapel Hill Writing Center. Literature Reviews. https://writingcenter.unc.edu/tips-and-tools/literature-reviews/ [Accessed September 8, 2022]
- Pautasso M. Ten simple rules for writing a literature review. PLoS Comput Biol. 2013, 9. doi: 10.1371/journal.pcbi.1003149.
Q: Is research complete without a review of related literature?
A research project is usually considered incomplete without a proper review of related literature. The review of related literature is a crucial component of any research project as it provides context for the research question, identifies gaps in existing literature, and ensures novelty by avoiding duplication. It also helps inform research design and supports arguments, highlights the significance of a study, and demonstrates your knowledge an expertise.
Q: What is difference between RRL and RRS?
The key difference between an RRL and an RRS lies in their focus and scope. An RRL or review of related literature examines a broad range of literature, including theoretical frameworks, concepts, and empirical studies, to establish the context and significance of the research topic. On the other hand, an RRS or review of research studies specifically focuses on analyzing and summarizing previous research studies within a specific research domain to gain insights into methodologies, findings, and gaps in the existing body of knowledge. While there may be some overlap between the two, they serve distinct purposes and cover different aspects of the research process.
Q: Does review of related literature improve accuracy and validity of research?
Yes, a comprehensive review of related literature (RRL) plays a vital role in improving the accuracy and validity of research. It helps authors gain a deeper understanding and offers different perspectives on the research topic. RRL can help you identify research gaps, dictate the selection of appropriate research methodologies, enhance theoretical frameworks, avoid biases and errors, and even provide support for research design and interpretation. By building upon and critically engaging with existing related literature, researchers can ensure their work is rigorous, reliable, and contributes meaningfully to their field of study.
R Discovery is a literature search and research reading platform that accelerates your research discovery journey by keeping you updated on the latest, most relevant scholarly content. With 250M+ research articles sourced from trusted aggregators like CrossRef, Unpaywall, PubMed, PubMed Central, Open Alex and top publishing houses like Springer Nature, JAMA, IOP, Taylor & Francis, NEJM, BMJ, Karger, SAGE, Emerald Publishing and more, R Discovery puts a world of research at your fingertips.
Try R Discovery Prime FREE for 1 week or upgrade at just US$72 a year to access premium features that let you listen to research on the go, read in your language, collaborate with peers, auto sync with reference managers, and much more. Choose a simpler, smarter way to find and read research – Download the app and start your free 7-day trial today !
Related Posts

What are Experimental Groups in Research

What is IMRaD Format in Research?
Have a language expert improve your writing
Run a free plagiarism check in 10 minutes, generate accurate citations for free.
- Knowledge Base
Methodology
- How to Write a Literature Review | Guide, Examples, & Templates
How to Write a Literature Review | Guide, Examples, & Templates
Published on January 2, 2023 by Shona McCombes . Revised on September 11, 2023.
What is a literature review? A literature review is a survey of scholarly sources on a specific topic. It provides an overview of current knowledge, allowing you to identify relevant theories, methods, and gaps in the existing research that you can later apply to your paper, thesis, or dissertation topic .
There are five key steps to writing a literature review:
- Search for relevant literature
- Evaluate sources
- Identify themes, debates, and gaps
- Outline the structure
- Write your literature review
A good literature review doesn’t just summarize sources—it analyzes, synthesizes , and critically evaluates to give a clear picture of the state of knowledge on the subject.
Instantly correct all language mistakes in your text
Upload your document to correct all your mistakes in minutes

Table of contents
What is the purpose of a literature review, examples of literature reviews, step 1 – search for relevant literature, step 2 – evaluate and select sources, step 3 – identify themes, debates, and gaps, step 4 – outline your literature review’s structure, step 5 – write your literature review, free lecture slides, other interesting articles, frequently asked questions, introduction.
- Quick Run-through
- Step 1 & 2
When you write a thesis , dissertation , or research paper , you will likely have to conduct a literature review to situate your research within existing knowledge. The literature review gives you a chance to:
- Demonstrate your familiarity with the topic and its scholarly context
- Develop a theoretical framework and methodology for your research
- Position your work in relation to other researchers and theorists
- Show how your research addresses a gap or contributes to a debate
- Evaluate the current state of research and demonstrate your knowledge of the scholarly debates around your topic.
Writing literature reviews is a particularly important skill if you want to apply for graduate school or pursue a career in research. We’ve written a step-by-step guide that you can follow below.

Receive feedback on language, structure, and formatting
Professional editors proofread and edit your paper by focusing on:
- Academic style
- Vague sentences
- Style consistency
See an example

Writing literature reviews can be quite challenging! A good starting point could be to look at some examples, depending on what kind of literature review you’d like to write.
- Example literature review #1: “Why Do People Migrate? A Review of the Theoretical Literature” ( Theoretical literature review about the development of economic migration theory from the 1950s to today.)
- Example literature review #2: “Literature review as a research methodology: An overview and guidelines” ( Methodological literature review about interdisciplinary knowledge acquisition and production.)
- Example literature review #3: “The Use of Technology in English Language Learning: A Literature Review” ( Thematic literature review about the effects of technology on language acquisition.)
- Example literature review #4: “Learners’ Listening Comprehension Difficulties in English Language Learning: A Literature Review” ( Chronological literature review about how the concept of listening skills has changed over time.)
You can also check out our templates with literature review examples and sample outlines at the links below.
Download Word doc Download Google doc
Before you begin searching for literature, you need a clearly defined topic .
If you are writing the literature review section of a dissertation or research paper, you will search for literature related to your research problem and questions .
Make a list of keywords
Start by creating a list of keywords related to your research question. Include each of the key concepts or variables you’re interested in, and list any synonyms and related terms. You can add to this list as you discover new keywords in the process of your literature search.
- Social media, Facebook, Instagram, Twitter, Snapchat, TikTok
- Body image, self-perception, self-esteem, mental health
- Generation Z, teenagers, adolescents, youth
Search for relevant sources
Use your keywords to begin searching for sources. Some useful databases to search for journals and articles include:
- Your university’s library catalogue
- Google Scholar
- Project Muse (humanities and social sciences)
- Medline (life sciences and biomedicine)
- EconLit (economics)
- Inspec (physics, engineering and computer science)
You can also use boolean operators to help narrow down your search.
Make sure to read the abstract to find out whether an article is relevant to your question. When you find a useful book or article, you can check the bibliography to find other relevant sources.
You likely won’t be able to read absolutely everything that has been written on your topic, so it will be necessary to evaluate which sources are most relevant to your research question.
For each publication, ask yourself:
- What question or problem is the author addressing?
- What are the key concepts and how are they defined?
- What are the key theories, models, and methods?
- Does the research use established frameworks or take an innovative approach?
- What are the results and conclusions of the study?
- How does the publication relate to other literature in the field? Does it confirm, add to, or challenge established knowledge?
- What are the strengths and weaknesses of the research?
Make sure the sources you use are credible , and make sure you read any landmark studies and major theories in your field of research.
You can use our template to summarize and evaluate sources you’re thinking about using. Click on either button below to download.
Take notes and cite your sources
As you read, you should also begin the writing process. Take notes that you can later incorporate into the text of your literature review.
It is important to keep track of your sources with citations to avoid plagiarism . It can be helpful to make an annotated bibliography , where you compile full citation information and write a paragraph of summary and analysis for each source. This helps you remember what you read and saves time later in the process.
Don't submit your assignments before you do this
The academic proofreading tool has been trained on 1000s of academic texts. Making it the most accurate and reliable proofreading tool for students. Free citation check included.

Try for free
To begin organizing your literature review’s argument and structure, be sure you understand the connections and relationships between the sources you’ve read. Based on your reading and notes, you can look for:
- Trends and patterns (in theory, method or results): do certain approaches become more or less popular over time?
- Themes: what questions or concepts recur across the literature?
- Debates, conflicts and contradictions: where do sources disagree?
- Pivotal publications: are there any influential theories or studies that changed the direction of the field?
- Gaps: what is missing from the literature? Are there weaknesses that need to be addressed?
This step will help you work out the structure of your literature review and (if applicable) show how your own research will contribute to existing knowledge.
- Most research has focused on young women.
- There is an increasing interest in the visual aspects of social media.
- But there is still a lack of robust research on highly visual platforms like Instagram and Snapchat—this is a gap that you could address in your own research.
There are various approaches to organizing the body of a literature review. Depending on the length of your literature review, you can combine several of these strategies (for example, your overall structure might be thematic, but each theme is discussed chronologically).
Chronological
The simplest approach is to trace the development of the topic over time. However, if you choose this strategy, be careful to avoid simply listing and summarizing sources in order.
Try to analyze patterns, turning points and key debates that have shaped the direction of the field. Give your interpretation of how and why certain developments occurred.
If you have found some recurring central themes, you can organize your literature review into subsections that address different aspects of the topic.
For example, if you are reviewing literature about inequalities in migrant health outcomes, key themes might include healthcare policy, language barriers, cultural attitudes, legal status, and economic access.
Methodological
If you draw your sources from different disciplines or fields that use a variety of research methods , you might want to compare the results and conclusions that emerge from different approaches. For example:
- Look at what results have emerged in qualitative versus quantitative research
- Discuss how the topic has been approached by empirical versus theoretical scholarship
- Divide the literature into sociological, historical, and cultural sources
Theoretical
A literature review is often the foundation for a theoretical framework . You can use it to discuss various theories, models, and definitions of key concepts.
You might argue for the relevance of a specific theoretical approach, or combine various theoretical concepts to create a framework for your research.
Like any other academic text , your literature review should have an introduction , a main body, and a conclusion . What you include in each depends on the objective of your literature review.
The introduction should clearly establish the focus and purpose of the literature review.
Depending on the length of your literature review, you might want to divide the body into subsections. You can use a subheading for each theme, time period, or methodological approach.
As you write, you can follow these tips:
- Summarize and synthesize: give an overview of the main points of each source and combine them into a coherent whole
- Analyze and interpret: don’t just paraphrase other researchers — add your own interpretations where possible, discussing the significance of findings in relation to the literature as a whole
- Critically evaluate: mention the strengths and weaknesses of your sources
- Write in well-structured paragraphs: use transition words and topic sentences to draw connections, comparisons and contrasts
In the conclusion, you should summarize the key findings you have taken from the literature and emphasize their significance.
When you’ve finished writing and revising your literature review, don’t forget to proofread thoroughly before submitting. Not a language expert? Check out Scribbr’s professional proofreading services !
This article has been adapted into lecture slides that you can use to teach your students about writing a literature review.
Scribbr slides are free to use, customize, and distribute for educational purposes.
Open Google Slides Download PowerPoint
If you want to know more about the research process , methodology , research bias , or statistics , make sure to check out some of our other articles with explanations and examples.
- Sampling methods
- Simple random sampling
- Stratified sampling
- Cluster sampling
- Likert scales
- Reproducibility
Statistics
- Null hypothesis
- Statistical power
- Probability distribution
- Effect size
- Poisson distribution
Research bias
- Optimism bias
- Cognitive bias
- Implicit bias
- Hawthorne effect
- Anchoring bias
- Explicit bias
A literature review is a survey of scholarly sources (such as books, journal articles, and theses) related to a specific topic or research question .
It is often written as part of a thesis, dissertation , or research paper , in order to situate your work in relation to existing knowledge.
There are several reasons to conduct a literature review at the beginning of a research project:
- To familiarize yourself with the current state of knowledge on your topic
- To ensure that you’re not just repeating what others have already done
- To identify gaps in knowledge and unresolved problems that your research can address
- To develop your theoretical framework and methodology
- To provide an overview of the key findings and debates on the topic
Writing the literature review shows your reader how your work relates to existing research and what new insights it will contribute.
The literature review usually comes near the beginning of your thesis or dissertation . After the introduction , it grounds your research in a scholarly field and leads directly to your theoretical framework or methodology .
A literature review is a survey of credible sources on a topic, often used in dissertations , theses, and research papers . Literature reviews give an overview of knowledge on a subject, helping you identify relevant theories and methods, as well as gaps in existing research. Literature reviews are set up similarly to other academic texts , with an introduction , a main body, and a conclusion .
An annotated bibliography is a list of source references that has a short description (called an annotation ) for each of the sources. It is often assigned as part of the research process for a paper .
Cite this Scribbr article
If you want to cite this source, you can copy and paste the citation or click the “Cite this Scribbr article” button to automatically add the citation to our free Citation Generator.
McCombes, S. (2023, September 11). How to Write a Literature Review | Guide, Examples, & Templates. Scribbr. Retrieved August 12, 2024, from https://www.scribbr.com/dissertation/literature-review/
Is this article helpful?

Shona McCombes
Other students also liked, what is a theoretical framework | guide to organizing, what is a research methodology | steps & tips, how to write a research proposal | examples & templates, get unlimited documents corrected.
✔ Free APA citation check included ✔ Unlimited document corrections ✔ Specialized in correcting academic texts
Libraries | Research Guides
Literature reviews, what is a literature review, learning more about how to do a literature review.
- Planning the Review
- The Research Question
- Choosing Where to Search
- Organizing the Review
- Writing the Review
A literature review is a review and synthesis of existing research on a topic or research question. A literature review is meant to analyze the scholarly literature, make connections across writings and identify strengths, weaknesses, trends, and missing conversations. A literature review should address different aspects of a topic as it relates to your research question. A literature review goes beyond a description or summary of the literature you have read.
- Sage Research Methods Core This link opens in a new window SAGE Research Methods supports research at all levels by providing material to guide users through every step of the research process. SAGE Research Methods is the ultimate methods library with more than 1000 books, reference works, journal articles, and instructional videos by world-leading academics from across the social sciences, including the largest collection of qualitative methods books available online from any scholarly publisher. – Publisher

- Next: Planning the Review >>
- Last Updated: Jul 8, 2024 11:22 AM
- URL: https://libguides.northwestern.edu/literaturereviews
- USC Libraries
- Research Guides
Organizing Your Social Sciences Research Paper
- 5. The Literature Review
- Purpose of Guide
- Design Flaws to Avoid
- Independent and Dependent Variables
- Glossary of Research Terms
- Reading Research Effectively
- Narrowing a Topic Idea
- Broadening a Topic Idea
- Extending the Timeliness of a Topic Idea
- Academic Writing Style
- Applying Critical Thinking
- Choosing a Title
- Making an Outline
- Paragraph Development
- Research Process Video Series
- Executive Summary
- The C.A.R.S. Model
- Background Information
- The Research Problem/Question
- Theoretical Framework
- Citation Tracking
- Content Alert Services
- Evaluating Sources
- Primary Sources
- Secondary Sources
- Tiertiary Sources
- Scholarly vs. Popular Publications
- Qualitative Methods
- Quantitative Methods
- Insiderness
- Using Non-Textual Elements
- Limitations of the Study
- Common Grammar Mistakes
- Writing Concisely
- Avoiding Plagiarism
- Footnotes or Endnotes?
- Further Readings
- Generative AI and Writing
- USC Libraries Tutorials and Other Guides
- Bibliography
A literature review surveys prior research published in books, scholarly articles, and any other sources relevant to a particular issue, area of research, or theory, and by so doing, provides a description, summary, and critical evaluation of these works in relation to the research problem being investigated. Literature reviews are designed to provide an overview of sources you have used in researching a particular topic and to demonstrate to your readers how your research fits within existing scholarship about the topic.
Fink, Arlene. Conducting Research Literature Reviews: From the Internet to Paper . Fourth edition. Thousand Oaks, CA: SAGE, 2014.
Importance of a Good Literature Review
A literature review may consist of simply a summary of key sources, but in the social sciences, a literature review usually has an organizational pattern and combines both summary and synthesis, often within specific conceptual categories . A summary is a recap of the important information of the source, but a synthesis is a re-organization, or a reshuffling, of that information in a way that informs how you are planning to investigate a research problem. The analytical features of a literature review might:
- Give a new interpretation of old material or combine new with old interpretations,
- Trace the intellectual progression of the field, including major debates,
- Depending on the situation, evaluate the sources and advise the reader on the most pertinent or relevant research, or
- Usually in the conclusion of a literature review, identify where gaps exist in how a problem has been researched to date.
Given this, the purpose of a literature review is to:
- Place each work in the context of its contribution to understanding the research problem being studied.
- Describe the relationship of each work to the others under consideration.
- Identify new ways to interpret prior research.
- Reveal any gaps that exist in the literature.
- Resolve conflicts amongst seemingly contradictory previous studies.
- Identify areas of prior scholarship to prevent duplication of effort.
- Point the way in fulfilling a need for additional research.
- Locate your own research within the context of existing literature [very important].
Fink, Arlene. Conducting Research Literature Reviews: From the Internet to Paper. 2nd ed. Thousand Oaks, CA: Sage, 2005; Hart, Chris. Doing a Literature Review: Releasing the Social Science Research Imagination . Thousand Oaks, CA: Sage Publications, 1998; Jesson, Jill. Doing Your Literature Review: Traditional and Systematic Techniques . Los Angeles, CA: SAGE, 2011; Knopf, Jeffrey W. "Doing a Literature Review." PS: Political Science and Politics 39 (January 2006): 127-132; Ridley, Diana. The Literature Review: A Step-by-Step Guide for Students . 2nd ed. Los Angeles, CA: SAGE, 2012.
Types of Literature Reviews
It is important to think of knowledge in a given field as consisting of three layers. First, there are the primary studies that researchers conduct and publish. Second are the reviews of those studies that summarize and offer new interpretations built from and often extending beyond the primary studies. Third, there are the perceptions, conclusions, opinion, and interpretations that are shared informally among scholars that become part of the body of epistemological traditions within the field.
In composing a literature review, it is important to note that it is often this third layer of knowledge that is cited as "true" even though it often has only a loose relationship to the primary studies and secondary literature reviews. Given this, while literature reviews are designed to provide an overview and synthesis of pertinent sources you have explored, there are a number of approaches you could adopt depending upon the type of analysis underpinning your study.
Argumentative Review This form examines literature selectively in order to support or refute an argument, deeply embedded assumption, or philosophical problem already established in the literature. The purpose is to develop a body of literature that establishes a contrarian viewpoint. Given the value-laden nature of some social science research [e.g., educational reform; immigration control], argumentative approaches to analyzing the literature can be a legitimate and important form of discourse. However, note that they can also introduce problems of bias when they are used to make summary claims of the sort found in systematic reviews [see below].
Integrative Review Considered a form of research that reviews, critiques, and synthesizes representative literature on a topic in an integrated way such that new frameworks and perspectives on the topic are generated. The body of literature includes all studies that address related or identical hypotheses or research problems. A well-done integrative review meets the same standards as primary research in regard to clarity, rigor, and replication. This is the most common form of review in the social sciences.
Historical Review Few things rest in isolation from historical precedent. Historical literature reviews focus on examining research throughout a period of time, often starting with the first time an issue, concept, theory, phenomena emerged in the literature, then tracing its evolution within the scholarship of a discipline. The purpose is to place research in a historical context to show familiarity with state-of-the-art developments and to identify the likely directions for future research.
Methodological Review A review does not always focus on what someone said [findings], but how they came about saying what they say [method of analysis]. Reviewing methods of analysis provides a framework of understanding at different levels [i.e. those of theory, substantive fields, research approaches, and data collection and analysis techniques], how researchers draw upon a wide variety of knowledge ranging from the conceptual level to practical documents for use in fieldwork in the areas of ontological and epistemological consideration, quantitative and qualitative integration, sampling, interviewing, data collection, and data analysis. This approach helps highlight ethical issues which you should be aware of and consider as you go through your own study.
Systematic Review This form consists of an overview of existing evidence pertinent to a clearly formulated research question, which uses pre-specified and standardized methods to identify and critically appraise relevant research, and to collect, report, and analyze data from the studies that are included in the review. The goal is to deliberately document, critically evaluate, and summarize scientifically all of the research about a clearly defined research problem . Typically it focuses on a very specific empirical question, often posed in a cause-and-effect form, such as "To what extent does A contribute to B?" This type of literature review is primarily applied to examining prior research studies in clinical medicine and allied health fields, but it is increasingly being used in the social sciences.
Theoretical Review The purpose of this form is to examine the corpus of theory that has accumulated in regard to an issue, concept, theory, phenomena. The theoretical literature review helps to establish what theories already exist, the relationships between them, to what degree the existing theories have been investigated, and to develop new hypotheses to be tested. Often this form is used to help establish a lack of appropriate theories or reveal that current theories are inadequate for explaining new or emerging research problems. The unit of analysis can focus on a theoretical concept or a whole theory or framework.
NOTE: Most often the literature review will incorporate some combination of types. For example, a review that examines literature supporting or refuting an argument, assumption, or philosophical problem related to the research problem will also need to include writing supported by sources that establish the history of these arguments in the literature.
Baumeister, Roy F. and Mark R. Leary. "Writing Narrative Literature Reviews." Review of General Psychology 1 (September 1997): 311-320; Mark R. Fink, Arlene. Conducting Research Literature Reviews: From the Internet to Paper . 2nd ed. Thousand Oaks, CA: Sage, 2005; Hart, Chris. Doing a Literature Review: Releasing the Social Science Research Imagination . Thousand Oaks, CA: Sage Publications, 1998; Kennedy, Mary M. "Defining a Literature." Educational Researcher 36 (April 2007): 139-147; Petticrew, Mark and Helen Roberts. Systematic Reviews in the Social Sciences: A Practical Guide . Malden, MA: Blackwell Publishers, 2006; Torracro, Richard. "Writing Integrative Literature Reviews: Guidelines and Examples." Human Resource Development Review 4 (September 2005): 356-367; Rocco, Tonette S. and Maria S. Plakhotnik. "Literature Reviews, Conceptual Frameworks, and Theoretical Frameworks: Terms, Functions, and Distinctions." Human Ressource Development Review 8 (March 2008): 120-130; Sutton, Anthea. Systematic Approaches to a Successful Literature Review . Los Angeles, CA: Sage Publications, 2016.
Structure and Writing Style
I. Thinking About Your Literature Review
The structure of a literature review should include the following in support of understanding the research problem :
- An overview of the subject, issue, or theory under consideration, along with the objectives of the literature review,
- Division of works under review into themes or categories [e.g. works that support a particular position, those against, and those offering alternative approaches entirely],
- An explanation of how each work is similar to and how it varies from the others,
- Conclusions as to which pieces are best considered in their argument, are most convincing of their opinions, and make the greatest contribution to the understanding and development of their area of research.
The critical evaluation of each work should consider :
- Provenance -- what are the author's credentials? Are the author's arguments supported by evidence [e.g. primary historical material, case studies, narratives, statistics, recent scientific findings]?
- Methodology -- were the techniques used to identify, gather, and analyze the data appropriate to addressing the research problem? Was the sample size appropriate? Were the results effectively interpreted and reported?
- Objectivity -- is the author's perspective even-handed or prejudicial? Is contrary data considered or is certain pertinent information ignored to prove the author's point?
- Persuasiveness -- which of the author's theses are most convincing or least convincing?
- Validity -- are the author's arguments and conclusions convincing? Does the work ultimately contribute in any significant way to an understanding of the subject?
II. Development of the Literature Review
Four Basic Stages of Writing 1. Problem formulation -- which topic or field is being examined and what are its component issues? 2. Literature search -- finding materials relevant to the subject being explored. 3. Data evaluation -- determining which literature makes a significant contribution to the understanding of the topic. 4. Analysis and interpretation -- discussing the findings and conclusions of pertinent literature.
Consider the following issues before writing the literature review: Clarify If your assignment is not specific about what form your literature review should take, seek clarification from your professor by asking these questions: 1. Roughly how many sources would be appropriate to include? 2. What types of sources should I review (books, journal articles, websites; scholarly versus popular sources)? 3. Should I summarize, synthesize, or critique sources by discussing a common theme or issue? 4. Should I evaluate the sources in any way beyond evaluating how they relate to understanding the research problem? 5. Should I provide subheadings and other background information, such as definitions and/or a history? Find Models Use the exercise of reviewing the literature to examine how authors in your discipline or area of interest have composed their literature review sections. Read them to get a sense of the types of themes you might want to look for in your own research or to identify ways to organize your final review. The bibliography or reference section of sources you've already read, such as required readings in the course syllabus, are also excellent entry points into your own research. Narrow the Topic The narrower your topic, the easier it will be to limit the number of sources you need to read in order to obtain a good survey of relevant resources. Your professor will probably not expect you to read everything that's available about the topic, but you'll make the act of reviewing easier if you first limit scope of the research problem. A good strategy is to begin by searching the USC Libraries Catalog for recent books about the topic and review the table of contents for chapters that focuses on specific issues. You can also review the indexes of books to find references to specific issues that can serve as the focus of your research. For example, a book surveying the history of the Israeli-Palestinian conflict may include a chapter on the role Egypt has played in mediating the conflict, or look in the index for the pages where Egypt is mentioned in the text. Consider Whether Your Sources are Current Some disciplines require that you use information that is as current as possible. This is particularly true in disciplines in medicine and the sciences where research conducted becomes obsolete very quickly as new discoveries are made. However, when writing a review in the social sciences, a survey of the history of the literature may be required. In other words, a complete understanding the research problem requires you to deliberately examine how knowledge and perspectives have changed over time. Sort through other current bibliographies or literature reviews in the field to get a sense of what your discipline expects. You can also use this method to explore what is considered by scholars to be a "hot topic" and what is not.
III. Ways to Organize Your Literature Review
Chronology of Events If your review follows the chronological method, you could write about the materials according to when they were published. This approach should only be followed if a clear path of research building on previous research can be identified and that these trends follow a clear chronological order of development. For example, a literature review that focuses on continuing research about the emergence of German economic power after the fall of the Soviet Union. By Publication Order your sources by publication chronology, then, only if the order demonstrates a more important trend. For instance, you could order a review of literature on environmental studies of brown fields if the progression revealed, for example, a change in the soil collection practices of the researchers who wrote and/or conducted the studies. Thematic [“conceptual categories”] A thematic literature review is the most common approach to summarizing prior research in the social and behavioral sciences. Thematic reviews are organized around a topic or issue, rather than the progression of time, although the progression of time may still be incorporated into a thematic review. For example, a review of the Internet’s impact on American presidential politics could focus on the development of online political satire. While the study focuses on one topic, the Internet’s impact on American presidential politics, it would still be organized chronologically reflecting technological developments in media. The difference in this example between a "chronological" and a "thematic" approach is what is emphasized the most: themes related to the role of the Internet in presidential politics. Note that more authentic thematic reviews tend to break away from chronological order. A review organized in this manner would shift between time periods within each section according to the point being made. Methodological A methodological approach focuses on the methods utilized by the researcher. For the Internet in American presidential politics project, one methodological approach would be to look at cultural differences between the portrayal of American presidents on American, British, and French websites. Or the review might focus on the fundraising impact of the Internet on a particular political party. A methodological scope will influence either the types of documents in the review or the way in which these documents are discussed.
Other Sections of Your Literature Review Once you've decided on the organizational method for your literature review, the sections you need to include in the paper should be easy to figure out because they arise from your organizational strategy. In other words, a chronological review would have subsections for each vital time period; a thematic review would have subtopics based upon factors that relate to the theme or issue. However, sometimes you may need to add additional sections that are necessary for your study, but do not fit in the organizational strategy of the body. What other sections you include in the body is up to you. However, only include what is necessary for the reader to locate your study within the larger scholarship about the research problem.
Here are examples of other sections, usually in the form of a single paragraph, you may need to include depending on the type of review you write:
- Current Situation : Information necessary to understand the current topic or focus of the literature review.
- Sources Used : Describes the methods and resources [e.g., databases] you used to identify the literature you reviewed.
- History : The chronological progression of the field, the research literature, or an idea that is necessary to understand the literature review, if the body of the literature review is not already a chronology.
- Selection Methods : Criteria you used to select (and perhaps exclude) sources in your literature review. For instance, you might explain that your review includes only peer-reviewed [i.e., scholarly] sources.
- Standards : Description of the way in which you present your information.
- Questions for Further Research : What questions about the field has the review sparked? How will you further your research as a result of the review?
IV. Writing Your Literature Review
Once you've settled on how to organize your literature review, you're ready to write each section. When writing your review, keep in mind these issues.
Use Evidence A literature review section is, in this sense, just like any other academic research paper. Your interpretation of the available sources must be backed up with evidence [citations] that demonstrates that what you are saying is valid. Be Selective Select only the most important points in each source to highlight in the review. The type of information you choose to mention should relate directly to the research problem, whether it is thematic, methodological, or chronological. Related items that provide additional information, but that are not key to understanding the research problem, can be included in a list of further readings . Use Quotes Sparingly Some short quotes are appropriate if you want to emphasize a point, or if what an author stated cannot be easily paraphrased. Sometimes you may need to quote certain terminology that was coined by the author, is not common knowledge, or taken directly from the study. Do not use extensive quotes as a substitute for using your own words in reviewing the literature. Summarize and Synthesize Remember to summarize and synthesize your sources within each thematic paragraph as well as throughout the review. Recapitulate important features of a research study, but then synthesize it by rephrasing the study's significance and relating it to your own work and the work of others. Keep Your Own Voice While the literature review presents others' ideas, your voice [the writer's] should remain front and center. For example, weave references to other sources into what you are writing but maintain your own voice by starting and ending the paragraph with your own ideas and wording. Use Caution When Paraphrasing When paraphrasing a source that is not your own, be sure to represent the author's information or opinions accurately and in your own words. Even when paraphrasing an author’s work, you still must provide a citation to that work.
V. Common Mistakes to Avoid
These are the most common mistakes made in reviewing social science research literature.
- Sources in your literature review do not clearly relate to the research problem;
- You do not take sufficient time to define and identify the most relevant sources to use in the literature review related to the research problem;
- Relies exclusively on secondary analytical sources rather than including relevant primary research studies or data;
- Uncritically accepts another researcher's findings and interpretations as valid, rather than examining critically all aspects of the research design and analysis;
- Does not describe the search procedures that were used in identifying the literature to review;
- Reports isolated statistical results rather than synthesizing them in chi-squared or meta-analytic methods; and,
- Only includes research that validates assumptions and does not consider contrary findings and alternative interpretations found in the literature.
Cook, Kathleen E. and Elise Murowchick. “Do Literature Review Skills Transfer from One Course to Another?” Psychology Learning and Teaching 13 (March 2014): 3-11; Fink, Arlene. Conducting Research Literature Reviews: From the Internet to Paper . 2nd ed. Thousand Oaks, CA: Sage, 2005; Hart, Chris. Doing a Literature Review: Releasing the Social Science Research Imagination . Thousand Oaks, CA: Sage Publications, 1998; Jesson, Jill. Doing Your Literature Review: Traditional and Systematic Techniques . London: SAGE, 2011; Literature Review Handout. Online Writing Center. Liberty University; Literature Reviews. The Writing Center. University of North Carolina; Onwuegbuzie, Anthony J. and Rebecca Frels. Seven Steps to a Comprehensive Literature Review: A Multimodal and Cultural Approach . Los Angeles, CA: SAGE, 2016; Ridley, Diana. The Literature Review: A Step-by-Step Guide for Students . 2nd ed. Los Angeles, CA: SAGE, 2012; Randolph, Justus J. “A Guide to Writing the Dissertation Literature Review." Practical Assessment, Research, and Evaluation. vol. 14, June 2009; Sutton, Anthea. Systematic Approaches to a Successful Literature Review . Los Angeles, CA: Sage Publications, 2016; Taylor, Dena. The Literature Review: A Few Tips On Conducting It. University College Writing Centre. University of Toronto; Writing a Literature Review. Academic Skills Centre. University of Canberra.
Writing Tip
Break Out of Your Disciplinary Box!
Thinking interdisciplinarily about a research problem can be a rewarding exercise in applying new ideas, theories, or concepts to an old problem. For example, what might cultural anthropologists say about the continuing conflict in the Middle East? In what ways might geographers view the need for better distribution of social service agencies in large cities than how social workers might study the issue? You don’t want to substitute a thorough review of core research literature in your discipline for studies conducted in other fields of study. However, particularly in the social sciences, thinking about research problems from multiple vectors is a key strategy for finding new solutions to a problem or gaining a new perspective. Consult with a librarian about identifying research databases in other disciplines; almost every field of study has at least one comprehensive database devoted to indexing its research literature.
Frodeman, Robert. The Oxford Handbook of Interdisciplinarity . New York: Oxford University Press, 2010.
Another Writing Tip
Don't Just Review for Content!
While conducting a review of the literature, maximize the time you devote to writing this part of your paper by thinking broadly about what you should be looking for and evaluating. Review not just what scholars are saying, but how are they saying it. Some questions to ask:
- How are they organizing their ideas?
- What methods have they used to study the problem?
- What theories have been used to explain, predict, or understand their research problem?
- What sources have they cited to support their conclusions?
- How have they used non-textual elements [e.g., charts, graphs, figures, etc.] to illustrate key points?
When you begin to write your literature review section, you'll be glad you dug deeper into how the research was designed and constructed because it establishes a means for developing more substantial analysis and interpretation of the research problem.
Hart, Chris. Doing a Literature Review: Releasing the Social Science Research Imagination . Thousand Oaks, CA: Sage Publications, 1 998.
Yet Another Writing Tip
When Do I Know I Can Stop Looking and Move On?
Here are several strategies you can utilize to assess whether you've thoroughly reviewed the literature:
- Look for repeating patterns in the research findings . If the same thing is being said, just by different people, then this likely demonstrates that the research problem has hit a conceptual dead end. At this point consider: Does your study extend current research? Does it forge a new path? Or, does is merely add more of the same thing being said?
- Look at sources the authors cite to in their work . If you begin to see the same researchers cited again and again, then this is often an indication that no new ideas have been generated to address the research problem.
- Search Google Scholar to identify who has subsequently cited leading scholars already identified in your literature review [see next sub-tab]. This is called citation tracking and there are a number of sources that can help you identify who has cited whom, particularly scholars from outside of your discipline. Here again, if the same authors are being cited again and again, this may indicate no new literature has been written on the topic.
Onwuegbuzie, Anthony J. and Rebecca Frels. Seven Steps to a Comprehensive Literature Review: A Multimodal and Cultural Approach . Los Angeles, CA: Sage, 2016; Sutton, Anthea. Systematic Approaches to a Successful Literature Review . Los Angeles, CA: Sage Publications, 2016.
- << Previous: Theoretical Framework
- Next: Citation Tracking >>
- Last Updated: Aug 13, 2024 12:57 PM
- URL: https://libguides.usc.edu/writingguide
Harvey Cushing/John Hay Whitney Medical Library
- Collections
- Research Help
YSN Doctoral Programs: Steps in Conducting a Literature Review
- Biomedical Databases
- Global (Public Health) Databases
- Soc. Sci., History, and Law Databases
- Grey Literature
- Trials Registers
- Data and Statistics
- Public Policy
- Google Tips
- Recommended Books
- Steps in Conducting a Literature Review
What is a literature review?
A literature review is an integrated analysis -- not just a summary-- of scholarly writings and other relevant evidence related directly to your research question. That is, it represents a synthesis of the evidence that provides background information on your topic and shows a association between the evidence and your research question.
A literature review may be a stand alone work or the introduction to a larger research paper, depending on the assignment. Rely heavily on the guidelines your instructor has given you.
Why is it important?
A literature review is important because it:
- Explains the background of research on a topic.
- Demonstrates why a topic is significant to a subject area.
- Discovers relationships between research studies/ideas.
- Identifies major themes, concepts, and researchers on a topic.
- Identifies critical gaps and points of disagreement.
- Discusses further research questions that logically come out of the previous studies.
APA7 Style resources

APA Style Blog - for those harder to find answers
1. Choose a topic. Define your research question.
Your literature review should be guided by your central research question. The literature represents background and research developments related to a specific research question, interpreted and analyzed by you in a synthesized way.
- Make sure your research question is not too broad or too narrow. Is it manageable?
- Begin writing down terms that are related to your question. These will be useful for searches later.
- If you have the opportunity, discuss your topic with your professor and your class mates.
2. Decide on the scope of your review
How many studies do you need to look at? How comprehensive should it be? How many years should it cover?
- This may depend on your assignment. How many sources does the assignment require?
3. Select the databases you will use to conduct your searches.
Make a list of the databases you will search.
Where to find databases:
- use the tabs on this guide
- Find other databases in the Nursing Information Resources web page
- More on the Medical Library web page
- ... and more on the Yale University Library web page
4. Conduct your searches to find the evidence. Keep track of your searches.
- Use the key words in your question, as well as synonyms for those words, as terms in your search. Use the database tutorials for help.
- Save the searches in the databases. This saves time when you want to redo, or modify, the searches. It is also helpful to use as a guide is the searches are not finding any useful results.
- Review the abstracts of research studies carefully. This will save you time.
- Use the bibliographies and references of research studies you find to locate others.
- Check with your professor, or a subject expert in the field, if you are missing any key works in the field.
- Ask your librarian for help at any time.
- Use a citation manager, such as EndNote as the repository for your citations. See the EndNote tutorials for help.
Review the literature
Some questions to help you analyze the research:
- What was the research question of the study you are reviewing? What were the authors trying to discover?
- Was the research funded by a source that could influence the findings?
- What were the research methodologies? Analyze its literature review, the samples and variables used, the results, and the conclusions.
- Does the research seem to be complete? Could it have been conducted more soundly? What further questions does it raise?
- If there are conflicting studies, why do you think that is?
- How are the authors viewed in the field? Has this study been cited? If so, how has it been analyzed?
Tips:
- Review the abstracts carefully.
- Keep careful notes so that you may track your thought processes during the research process.
- Create a matrix of the studies for easy analysis, and synthesis, across all of the studies.
- << Previous: Recommended Books
- Last Updated: Jun 20, 2024 9:08 AM
- URL: https://guides.library.yale.edu/YSNDoctoral
Research Methods
- Getting Started
- Literature Review Research
- Research Design
- Research Design By Discipline
- SAGE Research Methods
- Teaching with SAGE Research Methods
Literature Review
- What is a Literature Review?
- What is NOT a Literature Review?
- Purposes of a Literature Review
- Types of Literature Reviews
- Literature Reviews vs. Systematic Reviews
- Systematic vs. Meta-Analysis
Literature Review is a comprehensive survey of the works published in a particular field of study or line of research, usually over a specific period of time, in the form of an in-depth, critical bibliographic essay or annotated list in which attention is drawn to the most significant works.
Also, we can define a literature review as the collected body of scholarly works related to a topic:
- Summarizes and analyzes previous research relevant to a topic
- Includes scholarly books and articles published in academic journals
- Can be an specific scholarly paper or a section in a research paper
The objective of a Literature Review is to find previous published scholarly works relevant to an specific topic
- Help gather ideas or information
- Keep up to date in current trends and findings
- Help develop new questions
A literature review is important because it:
- Explains the background of research on a topic.
- Demonstrates why a topic is significant to a subject area.
- Helps focus your own research questions or problems
- Discovers relationships between research studies/ideas.
- Suggests unexplored ideas or populations
- Identifies major themes, concepts, and researchers on a topic.
- Tests assumptions; may help counter preconceived ideas and remove unconscious bias.
- Identifies critical gaps, points of disagreement, or potentially flawed methodology or theoretical approaches.
- Indicates potential directions for future research.
All content in this section is from Literature Review Research from Old Dominion University
Keep in mind the following, a literature review is NOT:
Not an essay
Not an annotated bibliography in which you summarize each article that you have reviewed. A literature review goes beyond basic summarizing to focus on the critical analysis of the reviewed works and their relationship to your research question.
Not a research paper where you select resources to support one side of an issue versus another. A lit review should explain and consider all sides of an argument in order to avoid bias, and areas of agreement and disagreement should be highlighted.
A literature review serves several purposes. For example, it
- provides thorough knowledge of previous studies; introduces seminal works.
- helps focus one’s own research topic.
- identifies a conceptual framework for one’s own research questions or problems; indicates potential directions for future research.
- suggests previously unused or underused methodologies, designs, quantitative and qualitative strategies.
- identifies gaps in previous studies; identifies flawed methodologies and/or theoretical approaches; avoids replication of mistakes.
- helps the researcher avoid repetition of earlier research.
- suggests unexplored populations.
- determines whether past studies agree or disagree; identifies controversy in the literature.
- tests assumptions; may help counter preconceived ideas and remove unconscious bias.
As Kennedy (2007) notes*, it is important to think of knowledge in a given field as consisting of three layers. First, there are the primary studies that researchers conduct and publish. Second are the reviews of those studies that summarize and offer new interpretations built from and often extending beyond the original studies. Third, there are the perceptions, conclusions, opinion, and interpretations that are shared informally that become part of the lore of field. In composing a literature review, it is important to note that it is often this third layer of knowledge that is cited as "true" even though it often has only a loose relationship to the primary studies and secondary literature reviews.
Given this, while literature reviews are designed to provide an overview and synthesis of pertinent sources you have explored, there are several approaches to how they can be done, depending upon the type of analysis underpinning your study. Listed below are definitions of types of literature reviews:
Argumentative Review This form examines literature selectively in order to support or refute an argument, deeply imbedded assumption, or philosophical problem already established in the literature. The purpose is to develop a body of literature that establishes a contrarian viewpoint. Given the value-laden nature of some social science research [e.g., educational reform; immigration control], argumentative approaches to analyzing the literature can be a legitimate and important form of discourse. However, note that they can also introduce problems of bias when they are used to to make summary claims of the sort found in systematic reviews.
Integrative Review Considered a form of research that reviews, critiques, and synthesizes representative literature on a topic in an integrated way such that new frameworks and perspectives on the topic are generated. The body of literature includes all studies that address related or identical hypotheses. A well-done integrative review meets the same standards as primary research in regard to clarity, rigor, and replication.
Historical Review Few things rest in isolation from historical precedent. Historical reviews are focused on examining research throughout a period of time, often starting with the first time an issue, concept, theory, phenomena emerged in the literature, then tracing its evolution within the scholarship of a discipline. The purpose is to place research in a historical context to show familiarity with state-of-the-art developments and to identify the likely directions for future research.
Methodological Review A review does not always focus on what someone said [content], but how they said it [method of analysis]. This approach provides a framework of understanding at different levels (i.e. those of theory, substantive fields, research approaches and data collection and analysis techniques), enables researchers to draw on a wide variety of knowledge ranging from the conceptual level to practical documents for use in fieldwork in the areas of ontological and epistemological consideration, quantitative and qualitative integration, sampling, interviewing, data collection and data analysis, and helps highlight many ethical issues which we should be aware of and consider as we go through our study.
Systematic Review This form consists of an overview of existing evidence pertinent to a clearly formulated research question, which uses pre-specified and standardized methods to identify and critically appraise relevant research, and to collect, report, and analyse data from the studies that are included in the review. Typically it focuses on a very specific empirical question, often posed in a cause-and-effect form, such as "To what extent does A contribute to B?"
Theoretical Review The purpose of this form is to concretely examine the corpus of theory that has accumulated in regard to an issue, concept, theory, phenomena. The theoretical literature review help establish what theories already exist, the relationships between them, to what degree the existing theories have been investigated, and to develop new hypotheses to be tested. Often this form is used to help establish a lack of appropriate theories or reveal that current theories are inadequate for explaining new or emerging research problems. The unit of analysis can focus on a theoretical concept or a whole theory or framework.
* Kennedy, Mary M. "Defining a Literature." Educational Researcher 36 (April 2007): 139-147.
All content in this section is from The Literature Review created by Dr. Robert Larabee USC
Robinson, P. and Lowe, J. (2015), Literature reviews vs systematic reviews. Australian and New Zealand Journal of Public Health, 39: 103-103. doi: 10.1111/1753-6405.12393

What's in the name? The difference between a Systematic Review and a Literature Review, and why it matters . By Lynn Kysh from University of Southern California

Systematic review or meta-analysis?
A systematic review answers a defined research question by collecting and summarizing all empirical evidence that fits pre-specified eligibility criteria.
A meta-analysis is the use of statistical methods to summarize the results of these studies.
Systematic reviews, just like other research articles, can be of varying quality. They are a significant piece of work (the Centre for Reviews and Dissemination at York estimates that a team will take 9-24 months), and to be useful to other researchers and practitioners they should have:
- clearly stated objectives with pre-defined eligibility criteria for studies
- explicit, reproducible methodology
- a systematic search that attempts to identify all studies
- assessment of the validity of the findings of the included studies (e.g. risk of bias)
- systematic presentation, and synthesis, of the characteristics and findings of the included studies
Not all systematic reviews contain meta-analysis.
Meta-analysis is the use of statistical methods to summarize the results of independent studies. By combining information from all relevant studies, meta-analysis can provide more precise estimates of the effects of health care than those derived from the individual studies included within a review. More information on meta-analyses can be found in Cochrane Handbook, Chapter 9 .
A meta-analysis goes beyond critique and integration and conducts secondary statistical analysis on the outcomes of similar studies. It is a systematic review that uses quantitative methods to synthesize and summarize the results.
An advantage of a meta-analysis is the ability to be completely objective in evaluating research findings. Not all topics, however, have sufficient research evidence to allow a meta-analysis to be conducted. In that case, an integrative review is an appropriate strategy.
Some of the content in this section is from Systematic reviews and meta-analyses: step by step guide created by Kate McAllister.
- << Previous: Getting Started
- Next: Research Design >>
- Last Updated: Jul 15, 2024 10:34 AM
- URL: https://guides.lib.udel.edu/researchmethods
- UConn Library
- Literature Review: The What, Why and How-to Guide
- Introduction
Literature Review: The What, Why and How-to Guide — Introduction
- Getting Started
- How to Pick a Topic
- Strategies to Find Sources
- Evaluating Sources & Lit. Reviews
- Tips for Writing Literature Reviews
- Writing Literature Review: Useful Sites
- Citation Resources
- Other Academic Writings
What are Literature Reviews?
So, what is a literature review? "A literature review is an account of what has been published on a topic by accredited scholars and researchers. In writing the literature review, your purpose is to convey to your reader what knowledge and ideas have been established on a topic, and what their strengths and weaknesses are. As a piece of writing, the literature review must be defined by a guiding concept (e.g., your research objective, the problem or issue you are discussing, or your argumentative thesis). It is not just a descriptive list of the material available, or a set of summaries." Taylor, D. The literature review: A few tips on conducting it . University of Toronto Health Sciences Writing Centre.
Goals of Literature Reviews
What are the goals of creating a Literature Review? A literature could be written to accomplish different aims:
- To develop a theory or evaluate an existing theory
- To summarize the historical or existing state of a research topic
- Identify a problem in a field of research
Baumeister, R. F., & Leary, M. R. (1997). Writing narrative literature reviews . Review of General Psychology , 1 (3), 311-320.
What kinds of sources require a Literature Review?
- A research paper assigned in a course
- A thesis or dissertation
- A grant proposal
- An article intended for publication in a journal
All these instances require you to collect what has been written about your research topic so that you can demonstrate how your own research sheds new light on the topic.
Types of Literature Reviews
What kinds of literature reviews are written?
Narrative review: The purpose of this type of review is to describe the current state of the research on a specific topic/research and to offer a critical analysis of the literature reviewed. Studies are grouped by research/theoretical categories, and themes and trends, strengths and weakness, and gaps are identified. The review ends with a conclusion section which summarizes the findings regarding the state of the research of the specific study, the gaps identify and if applicable, explains how the author's research will address gaps identify in the review and expand the knowledge on the topic reviewed.
- Example : Predictors and Outcomes of U.S. Quality Maternity Leave: A Review and Conceptual Framework: 10.1177/08948453211037398
Systematic review : "The authors of a systematic review use a specific procedure to search the research literature, select the studies to include in their review, and critically evaluate the studies they find." (p. 139). Nelson, L. K. (2013). Research in Communication Sciences and Disorders . Plural Publishing.
- Example : The effect of leave policies on increasing fertility: a systematic review: 10.1057/s41599-022-01270-w
Meta-analysis : "Meta-analysis is a method of reviewing research findings in a quantitative fashion by transforming the data from individual studies into what is called an effect size and then pooling and analyzing this information. The basic goal in meta-analysis is to explain why different outcomes have occurred in different studies." (p. 197). Roberts, M. C., & Ilardi, S. S. (2003). Handbook of Research Methods in Clinical Psychology . Blackwell Publishing.
- Example : Employment Instability and Fertility in Europe: A Meta-Analysis: 10.1215/00703370-9164737
Meta-synthesis : "Qualitative meta-synthesis is a type of qualitative study that uses as data the findings from other qualitative studies linked by the same or related topic." (p.312). Zimmer, L. (2006). Qualitative meta-synthesis: A question of dialoguing with texts . Journal of Advanced Nursing , 53 (3), 311-318.
- Example : Women’s perspectives on career successes and barriers: A qualitative meta-synthesis: 10.1177/05390184221113735
Literature Reviews in the Health Sciences
- UConn Health subject guide on systematic reviews Explanation of the different review types used in health sciences literature as well as tools to help you find the right review type
- << Previous: Getting Started
- Next: How to Pick a Topic >>
- Last Updated: Sep 21, 2022 2:16 PM
- URL: https://guides.lib.uconn.edu/literaturereview
- Maps & Floorplans
- Libraries A-Z

- Ellis Library (main)
- Engineering Library
- Geological Sciences
- Journalism Library
- Law Library
- Mathematical Sciences
- MU Digital Collections
- Veterinary Medical
- More Libraries...
- Instructional Services
- Course Reserves
- Course Guides
- Schedule a Library Class
- Class Assessment Forms
- Recordings & Tutorials
- Research & Writing Help
- More class resources
- Places to Study
- Borrow, Request & Renew
- Call Numbers
- Computers, Printers, Scanners & Software
- Digital Media Lab
- Equipment Lending: Laptops, cameras, etc.
- Subject Librarians
- Writing Tutors
- More In the Library...
- Undergraduate Students
- Graduate Students
- Faculty & Staff
- Researcher Support
- Distance Learners
- International Students
- More Services for...
- View my MU Libraries Account (login & click on My Library Account)
- View my MOBIUS Checkouts
- Renew my Books (login & click on My Loans)
- Place a Hold on a Book
- Request Books from Depository
- View my ILL@MU Account
- Set Up Alerts in Databases
- More Account Information...
Introduction to Literature Reviews
Introduction.
- Step One: Define
- Step Two: Research
- Step Three: Write
- Suggested Readings
A literature review is a written work that :
- Compiles significant research published on a topic by accredited scholars and researchers;
- Surveys scholarly articles, books, dissertations, conference proceedings, and other sources;
- Examines contrasting perspectives, theoretical approaches, methodologies, findings, results, conclusions.
- Reviews critically, analyzes, and synthesizes existing research on a topic; and,
- Performs a thorough “re” view, “overview”, or “look again” of past and current works on a subject, issue, or theory.
From these analyses, the writer then offers an overview of the current status of a particular area of knowledge from both a practical and theoretical perspective.
Literature reviews are important because they are usually a required step in a thesis proposal (Master's or PhD). The proposal will not be well-supported without a literature review. Also, literature reviews are important because they help you learn important authors and ideas in your field. This is useful for your coursework and your writing. Knowing key authors also helps you become acquainted with other researchers in your field.
Look at this diagram and imagine that your research is the "something new." This shows how your research should relate to major works and other sources.
Olivia Whitfield | Graduate Reference Assistant | 2012-2015
- Next: Step One: Define >>
- Last Updated: Jul 9, 2024 9:53 AM
- URL: https://libraryguides.missouri.edu/literaturereview


- University of Texas Libraries
Literature Reviews
- What is a literature review?
- Steps in the Literature Review Process
- Define your research question
- Determine inclusion and exclusion criteria
- Choose databases and search
- Review Results
- Synthesize Results
- Analyze Results
- Librarian Support
- Artificial Intelligence (AI) Tools
What is a Literature Review?
A literature or narrative review is a comprehensive review and analysis of the published literature on a specific topic or research question. The literature that is reviewed contains: books, articles, academic articles, conference proceedings, association papers, and dissertations. It contains the most pertinent studies and points to important past and current research and practices. It provides background and context, and shows how your research will contribute to the field.
A literature review should:
- Provide a comprehensive and updated review of the literature;
- Explain why this review has taken place;
- Articulate a position or hypothesis;
- Acknowledge and account for conflicting and corroborating points of view
From S age Research Methods
Purpose of a Literature Review
A literature review can be written as an introduction to a study to:
- Demonstrate how a study fills a gap in research
- Compare a study with other research that's been done
Or it can be a separate work (a research article on its own) which:
- Organizes or describes a topic
- Describes variables within a particular issue/problem
Limitations of a Literature Review
Some of the limitations of a literature review are:
- It's a snapshot in time. Unlike other reviews, this one has beginning, a middle and an end. There may be future developments that could make your work less relevant.
- It may be too focused. Some niche studies may miss the bigger picture.
- It can be difficult to be comprehensive. There is no way to make sure all the literature on a topic was considered.
- It is easy to be biased if you stick to top tier journals. There may be other places where people are publishing exemplary research. Look to open access publications and conferences to reflect a more inclusive collection. Also, make sure to include opposing views (and not just supporting evidence).
Source: Grant, Maria J., and Andrew Booth. “A Typology of Reviews: An Analysis of 14 Review Types and Associated Methodologies.” Health Information & Libraries Journal, vol. 26, no. 2, June 2009, pp. 91–108. Wiley Online Library, doi:10.1111/j.1471-1842.2009.00848.x.
Meryl Brodsky : Communication and Information Studies
Hannah Chapman Tripp : Biology, Neuroscience
Carolyn Cunningham : Human Development & Family Sciences, Psychology, Sociology
Larayne Dallas : Engineering
Janelle Hedstrom : Special Education, Curriculum & Instruction, Ed Leadership & Policy
Susan Macicak : Linguistics
Imelda Vetter : Dell Medical School
For help in other subject areas, please see the guide to library specialists by subject .
Periodically, UT Libraries runs a workshop covering the basics and library support for literature reviews. While we try to offer these once per academic year, we find providing the recording to be helpful to community members who have missed the session. Following is the most recent recording of the workshop, Conducting a Literature Review. To view the recording, a UT login is required.
- October 26, 2022 recording
- Last Updated: Aug 13, 2024 1:52 PM
- URL: https://guides.lib.utexas.edu/literaturereviews

University Libraries
Literature review.
- What is a Literature Review?
- What is Its Purpose?
- 1. Select a Topic
- 2. Set the Topic in Context
- 3. Types of Information Sources
- 4. Use Information Sources
- 5. Get the Information
- 6. Organize / Manage the Information
- 7. Position the Literature Review
- 8. Write the Literature Review

A literature review is a comprehensive summary of previous research on a topic. The literature review surveys scholarly articles, books, and other sources relevant to a particular area of research. The review should enumerate, describe, summarize, objectively evaluate and clarify this previous research. It should give a theoretical base for the research and help you (the author) determine the nature of your research. The literature review acknowledges the work of previous researchers, and in so doing, assures the reader that your work has been well conceived. It is assumed that by mentioning a previous work in the field of study, that the author has read, evaluated, and assimiliated that work into the work at hand.
A literature review creates a "landscape" for the reader, giving her or him a full understanding of the developments in the field. This landscape informs the reader that the author has indeed assimilated all (or the vast majority of) previous, significant works in the field into her or his research.
"In writing the literature review, the purpose is to convey to the reader what knowledge and ideas have been established on a topic, and what their strengths and weaknesses are. The literature review must be defined by a guiding concept (eg. your research objective, the problem or issue you are discussing, or your argumentative thesis). It is not just a descriptive list of the material available, or a set of summaries.( http://www.writing.utoronto.ca/advice/specific-types-of-writing/literature-review )
Recommended Reading

- Next: What is Its Purpose? >>
- Last Updated: Oct 2, 2023 12:34 PM
What is a Literature Review? How to Write It (with Examples)

A literature review is a critical analysis and synthesis of existing research on a particular topic. It provides an overview of the current state of knowledge, identifies gaps, and highlights key findings in the literature. 1 The purpose of a literature review is to situate your own research within the context of existing scholarship, demonstrating your understanding of the topic and showing how your work contributes to the ongoing conversation in the field. Learning how to write a literature review is a critical tool for successful research. Your ability to summarize and synthesize prior research pertaining to a certain topic demonstrates your grasp on the topic of study, and assists in the learning process.
Table of Contents
- What is the purpose of literature review?
- a. Habitat Loss and Species Extinction:
- b. Range Shifts and Phenological Changes:
- c. Ocean Acidification and Coral Reefs:
- d. Adaptive Strategies and Conservation Efforts:
How to write a good literature review
- Choose a Topic and Define the Research Question:
- Decide on the Scope of Your Review:
- Select Databases for Searches:
- Conduct Searches and Keep Track:
- Review the Literature:
- Organize and Write Your Literature Review:
- How to write a literature review faster with Paperpal?
- Frequently asked questions
What is a literature review?
A well-conducted literature review demonstrates the researcher’s familiarity with the existing literature, establishes the context for their own research, and contributes to scholarly conversations on the topic. One of the purposes of a literature review is also to help researchers avoid duplicating previous work and ensure that their research is informed by and builds upon the existing body of knowledge.

What is the purpose of literature review?
A literature review serves several important purposes within academic and research contexts. Here are some key objectives and functions of a literature review: 2
1. Contextualizing the Research Problem: The literature review provides a background and context for the research problem under investigation. It helps to situate the study within the existing body of knowledge.
2. Identifying Gaps in Knowledge: By identifying gaps, contradictions, or areas requiring further research, the researcher can shape the research question and justify the significance of the study. This is crucial for ensuring that the new research contributes something novel to the field.
Find academic papers related to your research topic faster. Try Research on Paperpal
3. Understanding Theoretical and Conceptual Frameworks: Literature reviews help researchers gain an understanding of the theoretical and conceptual frameworks used in previous studies. This aids in the development of a theoretical framework for the current research.
4. Providing Methodological Insights: Another purpose of literature reviews is that it allows researchers to learn about the methodologies employed in previous studies. This can help in choosing appropriate research methods for the current study and avoiding pitfalls that others may have encountered.
5. Establishing Credibility: A well-conducted literature review demonstrates the researcher’s familiarity with existing scholarship, establishing their credibility and expertise in the field. It also helps in building a solid foundation for the new research.
6. Informing Hypotheses or Research Questions: The literature review guides the formulation of hypotheses or research questions by highlighting relevant findings and areas of uncertainty in existing literature.
Literature review example
Let’s delve deeper with a literature review example: Let’s say your literature review is about the impact of climate change on biodiversity. You might format your literature review into sections such as the effects of climate change on habitat loss and species extinction, phenological changes, and marine biodiversity. Each section would then summarize and analyze relevant studies in those areas, highlighting key findings and identifying gaps in the research. The review would conclude by emphasizing the need for further research on specific aspects of the relationship between climate change and biodiversity. The following literature review template provides a glimpse into the recommended literature review structure and content, demonstrating how research findings are organized around specific themes within a broader topic.
Literature Review on Climate Change Impacts on Biodiversity:
Climate change is a global phenomenon with far-reaching consequences, including significant impacts on biodiversity. This literature review synthesizes key findings from various studies:
a. Habitat Loss and Species Extinction:
Climate change-induced alterations in temperature and precipitation patterns contribute to habitat loss, affecting numerous species (Thomas et al., 2004). The review discusses how these changes increase the risk of extinction, particularly for species with specific habitat requirements.
b. Range Shifts and Phenological Changes:
Observations of range shifts and changes in the timing of biological events (phenology) are documented in response to changing climatic conditions (Parmesan & Yohe, 2003). These shifts affect ecosystems and may lead to mismatches between species and their resources.
c. Ocean Acidification and Coral Reefs:
The review explores the impact of climate change on marine biodiversity, emphasizing ocean acidification’s threat to coral reefs (Hoegh-Guldberg et al., 2007). Changes in pH levels negatively affect coral calcification, disrupting the delicate balance of marine ecosystems.
d. Adaptive Strategies and Conservation Efforts:
Recognizing the urgency of the situation, the literature review discusses various adaptive strategies adopted by species and conservation efforts aimed at mitigating the impacts of climate change on biodiversity (Hannah et al., 2007). It emphasizes the importance of interdisciplinary approaches for effective conservation planning.

Strengthen your literature review with factual insights. Try Research on Paperpal for free!
Writing a literature review involves summarizing and synthesizing existing research on a particular topic. A good literature review format should include the following elements.
Introduction: The introduction sets the stage for your literature review, providing context and introducing the main focus of your review.
- Opening Statement: Begin with a general statement about the broader topic and its significance in the field.
- Scope and Purpose: Clearly define the scope of your literature review. Explain the specific research question or objective you aim to address.
- Organizational Framework: Briefly outline the structure of your literature review, indicating how you will categorize and discuss the existing research.
- Significance of the Study: Highlight why your literature review is important and how it contributes to the understanding of the chosen topic.
- Thesis Statement: Conclude the introduction with a concise thesis statement that outlines the main argument or perspective you will develop in the body of the literature review.
Body: The body of the literature review is where you provide a comprehensive analysis of existing literature, grouping studies based on themes, methodologies, or other relevant criteria.
- Organize by Theme or Concept: Group studies that share common themes, concepts, or methodologies. Discuss each theme or concept in detail, summarizing key findings and identifying gaps or areas of disagreement.
- Critical Analysis: Evaluate the strengths and weaknesses of each study. Discuss the methodologies used, the quality of evidence, and the overall contribution of each work to the understanding of the topic.
- Synthesis of Findings: Synthesize the information from different studies to highlight trends, patterns, or areas of consensus in the literature.
- Identification of Gaps: Discuss any gaps or limitations in the existing research and explain how your review contributes to filling these gaps.
- Transition between Sections: Provide smooth transitions between different themes or concepts to maintain the flow of your literature review.
Write and Cite as you go with Paperpal Research. Start now for free.
Conclusion: The conclusion of your literature review should summarize the main findings, highlight the contributions of the review, and suggest avenues for future research.
- Summary of Key Findings: Recap the main findings from the literature and restate how they contribute to your research question or objective.
- Contributions to the Field: Discuss the overall contribution of your literature review to the existing knowledge in the field.
- Implications and Applications: Explore the practical implications of the findings and suggest how they might impact future research or practice.
- Recommendations for Future Research: Identify areas that require further investigation and propose potential directions for future research in the field.
- Final Thoughts: Conclude with a final reflection on the importance of your literature review and its relevance to the broader academic community.

Conducting a literature review
Conducting a literature review is an essential step in research that involves reviewing and analyzing existing literature on a specific topic. It’s important to know how to do a literature review effectively, so here are the steps to follow: 1
Choose a Topic and Define the Research Question:
- Select a topic that is relevant to your field of study.
- Clearly define your research question or objective. Determine what specific aspect of the topic do you want to explore?
Decide on the Scope of Your Review:
- Determine the timeframe for your literature review. Are you focusing on recent developments, or do you want a historical overview?
- Consider the geographical scope. Is your review global, or are you focusing on a specific region?
- Define the inclusion and exclusion criteria. What types of sources will you include? Are there specific types of studies or publications you will exclude?
Select Databases for Searches:
- Identify relevant databases for your field. Examples include PubMed, IEEE Xplore, Scopus, Web of Science, and Google Scholar.
- Consider searching in library catalogs, institutional repositories, and specialized databases related to your topic.
Conduct Searches and Keep Track:
- Develop a systematic search strategy using keywords, Boolean operators (AND, OR, NOT), and other search techniques.
- Record and document your search strategy for transparency and replicability.
- Keep track of the articles, including publication details, abstracts, and links. Use citation management tools like EndNote, Zotero, or Mendeley to organize your references.
Review the Literature:
- Evaluate the relevance and quality of each source. Consider the methodology, sample size, and results of studies.
- Organize the literature by themes or key concepts. Identify patterns, trends, and gaps in the existing research.
- Summarize key findings and arguments from each source. Compare and contrast different perspectives.
- Identify areas where there is a consensus in the literature and where there are conflicting opinions.
- Provide critical analysis and synthesis of the literature. What are the strengths and weaknesses of existing research?
Organize and Write Your Literature Review:
- Literature review outline should be based on themes, chronological order, or methodological approaches.
- Write a clear and coherent narrative that synthesizes the information gathered.
- Use proper citations for each source and ensure consistency in your citation style (APA, MLA, Chicago, etc.).
- Conclude your literature review by summarizing key findings, identifying gaps, and suggesting areas for future research.
Whether you’re exploring a new research field or finding new angles to develop an existing topic, sifting through hundreds of papers can take more time than you have to spare. But what if you could find science-backed insights with verified citations in seconds? That’s the power of Paperpal’s new Research feature!
How to write a literature review faster with Paperpal?
Paperpal, an AI writing assistant, integrates powerful academic search capabilities within its writing platform. With the Research feature, you get 100% factual insights, with citations backed by 250M+ verified research articles, directly within your writing interface with the option to save relevant references in your Citation Library. By eliminating the need to switch tabs to find answers to all your research questions, Paperpal saves time and helps you stay focused on your writing.
Here’s how to use the Research feature:
- Ask a question: Get started with a new document on paperpal.com. Click on the “Research” feature and type your question in plain English. Paperpal will scour over 250 million research articles, including conference papers and preprints, to provide you with accurate insights and citations.
- Review and Save: Paperpal summarizes the information, while citing sources and listing relevant reads. You can quickly scan the results to identify relevant references and save these directly to your built-in citations library for later access.
- Cite with Confidence: Paperpal makes it easy to incorporate relevant citations and references into your writing, ensuring your arguments are well-supported by credible sources. This translates to a polished, well-researched literature review.
The literature review sample and detailed advice on writing and conducting a review will help you produce a well-structured report. But remember that a good literature review is an ongoing process, and it may be necessary to revisit and update it as your research progresses. By combining effortless research with an easy citation process, Paperpal Research streamlines the literature review process and empowers you to write faster and with more confidence. Try Paperpal Research now and see for yourself.
Frequently asked questions
A literature review is a critical and comprehensive analysis of existing literature (published and unpublished works) on a specific topic or research question and provides a synthesis of the current state of knowledge in a particular field. A well-conducted literature review is crucial for researchers to build upon existing knowledge, avoid duplication of efforts, and contribute to the advancement of their field. It also helps researchers situate their work within a broader context and facilitates the development of a sound theoretical and conceptual framework for their studies.
Literature review is a crucial component of research writing, providing a solid background for a research paper’s investigation. The aim is to keep professionals up to date by providing an understanding of ongoing developments within a specific field, including research methods, and experimental techniques used in that field, and present that knowledge in the form of a written report. Also, the depth and breadth of the literature review emphasizes the credibility of the scholar in his or her field.
Before writing a literature review, it’s essential to undertake several preparatory steps to ensure that your review is well-researched, organized, and focused. This includes choosing a topic of general interest to you and doing exploratory research on that topic, writing an annotated bibliography, and noting major points, especially those that relate to the position you have taken on the topic.
Literature reviews and academic research papers are essential components of scholarly work but serve different purposes within the academic realm. 3 A literature review aims to provide a foundation for understanding the current state of research on a particular topic, identify gaps or controversies, and lay the groundwork for future research. Therefore, it draws heavily from existing academic sources, including books, journal articles, and other scholarly publications. In contrast, an academic research paper aims to present new knowledge, contribute to the academic discourse, and advance the understanding of a specific research question. Therefore, it involves a mix of existing literature (in the introduction and literature review sections) and original data or findings obtained through research methods.
Literature reviews are essential components of academic and research papers, and various strategies can be employed to conduct them effectively. If you want to know how to write a literature review for a research paper, here are four common approaches that are often used by researchers. Chronological Review: This strategy involves organizing the literature based on the chronological order of publication. It helps to trace the development of a topic over time, showing how ideas, theories, and research have evolved. Thematic Review: Thematic reviews focus on identifying and analyzing themes or topics that cut across different studies. Instead of organizing the literature chronologically, it is grouped by key themes or concepts, allowing for a comprehensive exploration of various aspects of the topic. Methodological Review: This strategy involves organizing the literature based on the research methods employed in different studies. It helps to highlight the strengths and weaknesses of various methodologies and allows the reader to evaluate the reliability and validity of the research findings. Theoretical Review: A theoretical review examines the literature based on the theoretical frameworks used in different studies. This approach helps to identify the key theories that have been applied to the topic and assess their contributions to the understanding of the subject. It’s important to note that these strategies are not mutually exclusive, and a literature review may combine elements of more than one approach. The choice of strategy depends on the research question, the nature of the literature available, and the goals of the review. Additionally, other strategies, such as integrative reviews or systematic reviews, may be employed depending on the specific requirements of the research.
The literature review format can vary depending on the specific publication guidelines. However, there are some common elements and structures that are often followed. Here is a general guideline for the format of a literature review: Introduction: Provide an overview of the topic. Define the scope and purpose of the literature review. State the research question or objective. Body: Organize the literature by themes, concepts, or chronology. Critically analyze and evaluate each source. Discuss the strengths and weaknesses of the studies. Highlight any methodological limitations or biases. Identify patterns, connections, or contradictions in the existing research. Conclusion: Summarize the key points discussed in the literature review. Highlight the research gap. Address the research question or objective stated in the introduction. Highlight the contributions of the review and suggest directions for future research.
Both annotated bibliographies and literature reviews involve the examination of scholarly sources. While annotated bibliographies focus on individual sources with brief annotations, literature reviews provide a more in-depth, integrated, and comprehensive analysis of existing literature on a specific topic. The key differences are as follows:
| Annotated Bibliography | Literature Review | |
| Purpose | List of citations of books, articles, and other sources with a brief description (annotation) of each source. | Comprehensive and critical analysis of existing literature on a specific topic. |
| Focus | Summary and evaluation of each source, including its relevance, methodology, and key findings. | Provides an overview of the current state of knowledge on a particular subject and identifies gaps, trends, and patterns in existing literature. |
| Structure | Each citation is followed by a concise paragraph (annotation) that describes the source’s content, methodology, and its contribution to the topic. | The literature review is organized thematically or chronologically and involves a synthesis of the findings from different sources to build a narrative or argument. |
| Length | Typically 100-200 words | Length of literature review ranges from a few pages to several chapters |
| Independence | Each source is treated separately, with less emphasis on synthesizing the information across sources. | The writer synthesizes information from multiple sources to present a cohesive overview of the topic. |
References
- Denney, A. S., & Tewksbury, R. (2013). How to write a literature review. Journal of criminal justice education , 24 (2), 218-234.
- Pan, M. L. (2016). Preparing literature reviews: Qualitative and quantitative approaches . Taylor & Francis.
- Cantero, C. (2019). How to write a literature review. San José State University Writing Center .
Paperpal is an AI writing assistant that help academics write better, faster with real-time suggestions for in-depth language and grammar correction. Trained on millions of research manuscripts enhanced by professional academic editors, Paperpal delivers human precision at machine speed.
Try it for free or upgrade to Paperpal Prime , which unlocks unlimited access to premium features like academic translation, paraphrasing, contextual synonyms, consistency checks and more. It’s like always having a professional academic editor by your side! Go beyond limitations and experience the future of academic writing. Get Paperpal Prime now at just US$19 a month!
Related Reads:
- Empirical Research: A Comprehensive Guide for Academics
- How to Write a Scientific Paper in 10 Steps
- How Long Should a Chapter Be?
- How to Use Paperpal to Generate Emails & Cover Letters?
6 Tips for Post-Doc Researchers to Take Their Career to the Next Level
Self-plagiarism in research: what it is and how to avoid it, you may also like, academic integrity vs academic dishonesty: types & examples, dissertation printing and binding | types & comparison , what is a dissertation preface definition and examples , the ai revolution: authors’ role in upholding academic..., the future of academia: how ai tools are..., how to write a research proposal: (with examples..., how to write your research paper in apa..., how to choose a dissertation topic, how to write a phd research proposal, how to write an academic paragraph (step-by-step guide).
Review of Related Literature: Format, Example, & How to Make RRL
A review of related literature is a separate paper or a part of an article that collects and synthesizes discussion on a topic. Its purpose is to show the current state of research on the issue and highlight gaps in existing knowledge. A literature review can be included in a research paper or scholarly article, typically following the introduction and before the research methods section.

This article will clarify the definition, significance, and structure of a review of related literature. You’ll also learn how to organize your literature review and discover ideas for an RRL in different subjects.
🔤 What Is RRL?
- ❗ Significance of Literature Review
- 🔎 How to Search for Literature
- 🧩 Literature Review Structure
- 📋 Format of RRL — APA, MLA, & Others
- ✍️ How to Write an RRL
- 📚 Examples of RRL
🔗 References
A review of related literature (RRL) is a part of the research report that examines significant studies, theories, and concepts published in scholarly sources on a particular topic. An RRL includes 3 main components:
- A short overview and critique of the previous research.
- Similarities and differences between past studies and the current one.
- An explanation of the theoretical frameworks underpinning the research.
❗ Significance of Review of Related Literature
Although the goal of a review of related literature differs depending on the discipline and its intended use, its significance cannot be overstated. Here are some examples of how a review might be beneficial:
- It helps determine knowledge gaps .
- It saves from duplicating research that has already been conducted.
- It provides an overview of various research areas within the discipline.
- It demonstrates the researcher’s familiarity with the topic.
🔎 How to Perform a Literature Search
Including a description of your search strategy in the literature review section can significantly increase your grade. You can search sources with the following steps:
| You should specify all the keywords and their synonyms used to look for relevant sources. | |
| Using your search terms, look through the online (libraries and databases) and offline (books and journals) sources related to your topic. | |
| It is not possible to discuss all of the sources you have discovered. Instead, use the works of the most notable researchers and authors. | |
| From the remaining references, you should pick those with the most significant contribution to the research area development. | |
| Your literature should prioritize new publications over older ones to cover the latest research advancements. |
🧩 Literature Review Structure Example
The majority of literature reviews follow a standard introduction-body-conclusion structure. Let’s look at the RRL structure in detail.

Introduction of Review of Related Literature: Sample
An introduction should clarify the study topic and the depth of the information to be delivered. It should also explain the types of sources used. If your lit. review is part of a larger research proposal or project, you can combine its introductory paragraph with the introduction of your paper.
Here is a sample introduction to an RRL about cyberbullying:
Bullying has troubled people since the beginning of time. However, with modern technological advancements, especially social media, bullying has evolved into cyberbullying. As a result, nowadays, teenagers and adults cannot flee their bullies, which makes them feel lonely and helpless. This literature review will examine recent studies on cyberbullying.
Sample Review of Related Literature Thesis
A thesis statement should include the central idea of your literature review and the primary supporting elements you discovered in the literature. Thesis statements are typically put at the end of the introductory paragraph.
Look at a sample thesis of a review of related literature:
This literature review shows that scholars have recently covered the issues of bullies’ motivation, the impact of bullying on victims and aggressors, common cyberbullying techniques, and victims’ coping strategies. However, there is still no agreement on the best practices to address cyberbullying.
Literature Review Body Paragraph Example
The main body of a literature review should provide an overview of the existing research on the issue. Body paragraphs should not just summarize each source but analyze them. You can organize your paragraphs with these 3 elements:
- Claim . Start with a topic sentence linked to your literature review purpose.
- Evidence . Cite relevant information from your chosen sources.
- Discussion . Explain how the cited data supports your claim.
Here’s a literature review body paragraph example:
Scholars have examined the link between the aggressor and the victim. Beran et al. (2007) state that students bullied online often become cyberbullies themselves. Faucher et al. (2014) confirm this with their findings: they discovered that male and female students began engaging in cyberbullying after being subject to bullying. Hence, one can conclude that being a victim of bullying increases one’s likelihood of becoming a cyberbully.
Review of Related Literature: Conclusion
A conclusion presents a general consensus on the topic. Depending on your literature review purpose, it might include the following:
- Introduction to further research . If you write a literature review as part of a larger research project, you can present your research question in your conclusion .
- Overview of theories . You can summarize critical theories and concepts to help your reader understand the topic better.
- Discussion of the gap . If you identified a research gap in the reviewed literature, your conclusion could explain why that gap is significant.
Check out a conclusion example that discusses a research gap:
There is extensive research into bullies’ motivation, the consequences of bullying for victims and aggressors, strategies for bullying, and coping with it. Yet, scholars still have not reached a consensus on what to consider the best practices to combat cyberbullying. This question is of great importance because of the significant adverse effects of cyberbullying on victims and bullies.
📋 Format of RRL — APA, MLA, & Others
In this section, we will discuss how to format an RRL according to the most common citation styles: APA, Chicago, MLA, and Harvard.
Writing a literature review using the APA7 style requires the following text formatting:
| Times New Roman or Arial, 12 pt | |
| Double spacing | |
| All sides — 1″ (2.54 cm) | |
| Top right-hand corner, starting with the title page | |
- When using APA in-text citations , include the author’s last name and the year of publication in parentheses.
- For direct quotations , you must also add the page number. If you use sources without page numbers, such as websites or e-books, include a paragraph number instead.
- When referring to the author’s name in a sentence , you do not need to repeat it at the end of the sentence. Instead, include the year of publication inside the parentheses after their name.
- The reference list should be included at the end of your literature review. It is always alphabetized by the last name of the author (from A to Z), and the lines are indented one-half inch from the left margin of your paper. Do not forget to invert authors’ names (the last name should come first) and include the full titles of journals instead of their abbreviations. If you use an online source, add its URL.
The RRL format in the Chicago style is as follows:
| 12-pt Times New Roman, Arial, or Palatino | |
| Double spacing, single spacing is used to format block quotations, titles of tables and figures, footnotes, and bibliographical entries. | |
| All sides — 1″ (2.54 cm) | |
| Top right-hand corner. There should be no numbered pages on the title page or the page with the table of contents. |
- Author-date . You place your citations in brackets within the text, indicating the name of the author and the year of publication.
- Notes and bibliography . You place your citations in numbered footnotes or endnotes to connect the citation back to the source in the bibliography.
- The reference list, or bibliography , in Chicago style, is at the end of a literature review. The sources are arranged alphabetically and single-spaced. Each bibliography entry begins with the author’s name and the source’s title, followed by publication information, such as the city of publication, the publisher, and the year of publication.
Writing a literature review using the MLA style requires the following text formatting:
| Font | 12-pt Times New Roman or Arial |
| Line spacing | Double spacing |
| Margins | All sides — 1″ (2.54 cm) |
| Page numbers | Top right-hand corner. Your last name should precede the page number. |
| Title page | Not required. Instead, include a header in the top left-hand corner of the first page with content. It should contain: |
- In the MLA format, you can cite a source in the text by indicating the author’s last name and the page number in parentheses at the end of the citation. If the cited information takes several pages, you need to include all the page numbers.
- The reference list in MLA style is titled “ Works Cited .” In this section, all sources used in the paper should be listed in alphabetical order. Each entry should contain the author, title of the source, title of the journal or a larger volume, other contributors, version, number, publisher, and publication date.
The Harvard style requires you to use the following text formatting for your RRL:
| 12-pt Times New Roman or Arial | |
| Double spacing | |
| All sides — 1″ (2.54 cm) | |
| Top right-hand corner. Your last name should precede the page number. |
- In-text citations in the Harvard style include the author’s last name and the year of publication. If you are using a direct quote in your literature review, you need to add the page number as well.
- Arrange your list of references alphabetically. Each entry should contain the author’s last name, their initials, the year of publication, the title of the source, and other publication information, like the journal title and issue number or the publisher.
✍️ How to Write Review of Related Literature – Sample
Literature reviews can be organized in many ways depending on what you want to achieve with them. In this section, we will look at 3 examples of how you can write your RRL.

Thematic Literature Review
A thematic literature review is arranged around central themes or issues discussed in the sources. If you have identified some recurring themes in the literature, you can divide your RRL into sections that address various aspects of the topic. For example, if you examine studies on e-learning, you can distinguish such themes as the cost-effectiveness of online learning, the technologies used, and its effectiveness compared to traditional education.
Chronological Literature Review
A chronological literature review is a way to track the development of the topic over time. If you use this method, avoid merely listing and summarizing sources in chronological order. Instead, try to analyze the trends, turning moments, and critical debates that have shaped the field’s path. Also, you can give your interpretation of how and why specific advances occurred.
Methodological Literature Review
A methodological literature review differs from the preceding ones in that it usually doesn’t focus on the sources’ content. Instead, it is concerned with the research methods . So, if your references come from several disciplines or fields employing various research techniques, you can compare the findings and conclusions of different methodologies, for instance:
- empirical vs. theoretical studies;
- qualitative vs. quantitative research.
📚 Examples of Review of Related Literature and Studies
We have prepared a short example of RRL on climate change for you to see how everything works in practice!
Climate change is one of the most important issues nowadays. Based on a variety of facts, it is now clearer than ever that humans are altering the Earth's climate. The atmosphere and oceans have warmed, causing sea level rise, a significant loss of Arctic ice, and other climate-related changes. This literature review provides a thorough summary of research on climate change, focusing on climate change fingerprints and evidence of human influence on the Earth's climate system.
Physical Mechanisms and Evidence of Human Influence
Scientists are convinced that climate change is directly influenced by the emission of greenhouse gases. They have carefully analyzed various climate data and evidence, concluding that the majority of the observed global warming over the past 50 years cannot be explained by natural factors alone. Instead, there is compelling evidence pointing to a significant contribution of human activities, primarily the emission of greenhouse gases (Walker, 2014). For example, based on simple physics calculations, doubled carbon dioxide concentration in the atmosphere can lead to a global temperature increase of approximately 1 degree Celsius. (Elderfield, 2022). In order to determine the human influence on climate, scientists still have to analyze a lot of natural changes that affect temperature, precipitation, and other components of climate on timeframes ranging from days to decades and beyond.
Fingerprinting Climate Change
Fingerprinting climate change is a useful tool to identify the causes of global warming because different factors leave unique marks on climate records. This is evident when scientists look beyond overall temperature changes and examine how warming is distributed geographically and over time (Watson, 2022). By investigating these climate patterns, scientists can obtain a more complex understanding of the connections between natural climate variability and climate variability caused by human activity.
Modeling Climate Change and Feedback
To accurately predict the consequences of feedback mechanisms, the rate of warming, and regional climate change, scientists can employ sophisticated mathematical models of the atmosphere, ocean, land, and ice (the cryosphere). These models are grounded in well-established physical laws and incorporate the latest scientific understanding of climate-related processes (Shuckburgh, 2013). Although different climate models produce slightly varying projections for future warming, they all will agree that feedback mechanisms play a significant role in amplifying the initial warming caused by greenhouse gas emissions. (Meehl, 2019).
In conclusion, the literature on global warming indicates that there are well-understood physical processes that link variations in greenhouse gas concentrations to climate change. In addition, it covers the scientific proof that the rates of these gases in the atmosphere have increased and continue to rise fast. According to the sources, the majority of this recent change is almost definitely caused by greenhouse gas emissions produced by human activities. Citizens and governments can alter their energy production methods and consumption patterns to reduce greenhouse gas emissions and, thus, the magnitude of climate change. By acting now, society can prevent the worst consequences of climate change and build a more resilient and sustainable future for generations to come.
Have you ever struggled with finding the topic for an RRL in different subjects? Read the following paragraphs to get some ideas!
Nursing Literature Review Example
Many topics in the nursing field require research. For example, you can write a review of literature related to dengue fever . Give a general overview of dengue virus infections, including its clinical symptoms, diagnosis, prevention, and therapy.
Another good idea is to review related literature and studies about teenage pregnancy . This review can describe the effectiveness of specific programs for adolescent mothers and their children and summarize recommendations for preventing early pregnancy.
📝 Check out some more valuable examples below:
- Hospital Readmissions: Literature Review .
- Literature Review: Lower Sepsis Mortality Rates .
- Breast Cancer: Literature Review .
- Sexually Transmitted Diseases: Literature Review .
- PICO for Pressure Ulcers: Literature Review .
- COVID-19 Spread Prevention: Literature Review .
- Chronic Obstructive Pulmonary Disease: Literature Review .
- Hypertension Treatment Adherence: Literature Review .
- Neonatal Sepsis Prevention: Literature Review .
- Healthcare-Associated Infections: Literature Review .
- Understaffing in Nursing: Literature Review .
Psychology Literature Review Example
If you look for an RRL topic in psychology , you can write a review of related literature about stress . Summarize scientific evidence about stress stages, side effects, types, or reduction strategies. Or you can write a review of related literature about computer game addiction . In this case, you may concentrate on the neural mechanisms underlying the internet gaming disorder, compare it to other addictions, or evaluate treatment strategies.
A review of related literature about cyberbullying is another interesting option. You can highlight the impact of cyberbullying on undergraduate students’ academic, social, and emotional development.
📝 Look at the examples that we have prepared for you to come up with some more ideas:
- Mindfulness in Counseling: A Literature Review .
- Team-Building Across Cultures: Literature Review .
- Anxiety and Decision Making: Literature Review .
- Literature Review on Depression .
- Literature Review on Narcissism .
- Effects of Depression Among Adolescents .
- Causes and Effects of Anxiety in Children .
Literature Review — Sociology Example
Sociological research poses critical questions about social structures and phenomena. For example, you can write a review of related literature about child labor , exploring cultural beliefs and social norms that normalize the exploitation of children. Or you can create a review of related literature about social media . It can investigate the impact of social media on relationships between adolescents or the role of social networks on immigrants’ acculturation .
📝 You can find some more ideas below!
- Single Mothers’ Experiences of Relationships with Their Adolescent Sons .
- Teachers and Students’ Gender-Based Interactions .
- Gender Identity: Biological Perspective and Social Cognitive Theory .
- Gender: Culturally-Prescribed Role or Biological Sex .
- The Influence of Opioid Misuse on Academic Achievement of Veteran Students .
- The Importance of Ethics in Research .
- The Role of Family and Social Network Support in Mental Health .
Education Literature Review Example
For your education studies , you can write a review of related literature about academic performance to determine factors that affect student achievement and highlight research gaps. One more idea is to create a review of related literature on study habits , considering their role in the student’s life and academic outcomes.
You can also evaluate a computerized grading system in a review of related literature to single out its advantages and barriers to implementation. Or you can complete a review of related literature on instructional materials to identify their most common types and effects on student achievement.
📝 Find some inspiration in the examples below:
- Literature Review on Online Learning Challenges From COVID-19 .
- Education, Leadership, and Management: Literature Review .
- Literature Review: Standardized Testing Bias .
- Bullying of Disabled Children in School .
- Interventions and Letter & Sound Recognition: A Literature Review .
- Social-Emotional Skills Program for Preschoolers .
- Effectiveness of Educational Leadership Management Skills .
Business Research Literature Review
If you’re a business student, you can focus on customer satisfaction in your review of related literature. Discuss specific customer satisfaction features and how it is affected by service quality and prices. You can also create a theoretical literature review about consumer buying behavior to evaluate theories that have significantly contributed to understanding how consumers make purchasing decisions.
📝 Look at the examples to get more exciting ideas:
- Leadership and Communication: Literature Review .
- Human Resource Development: Literature Review .
- Project Management. Literature Review .
- Strategic HRM: A Literature Review .
- Customer Relationship Management: Literature Review .
- Literature Review on International Financial Reporting Standards .
- Cultures of Management: Literature Review .
To conclude, a review of related literature is a significant genre of scholarly works that can be applied in various disciplines and for multiple goals. The sources examined in an RRL provide theoretical frameworks for future studies and help create original research questions and hypotheses.
When you finish your outstanding literature review, don’t forget to check whether it sounds logical and coherent. Our text-to-speech tool can help you with that!
- Literature Reviews | University of North Carolina at Chapel Hill
- Writing a Literature Review | Purdue Online Writing Lab
- Learn How to Write a Review of Literature | University of Wisconsin-Madison
- The Literature Review: A Few Tips on Conducting It | University of Toronto
- Writing a Literature Review | UC San Diego
- Conduct a Literature Review | The University of Arizona
- Methods for Literature Reviews | National Library of Medicine
- Literature Reviews: 5. Write the Review | Georgia State University
How to Write an Animal Testing Essay: Tips for Argumentative & Persuasive Papers
Descriptive essay topics: examples, outline, & more.

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- PLoS Comput Biol
- v.9(7); 2013 Jul
Ten Simple Rules for Writing a Literature Review
Marco pautasso.
1 Centre for Functional and Evolutionary Ecology (CEFE), CNRS, Montpellier, France
2 Centre for Biodiversity Synthesis and Analysis (CESAB), FRB, Aix-en-Provence, France
Literature reviews are in great demand in most scientific fields. Their need stems from the ever-increasing output of scientific publications [1] . For example, compared to 1991, in 2008 three, eight, and forty times more papers were indexed in Web of Science on malaria, obesity, and biodiversity, respectively [2] . Given such mountains of papers, scientists cannot be expected to examine in detail every single new paper relevant to their interests [3] . Thus, it is both advantageous and necessary to rely on regular summaries of the recent literature. Although recognition for scientists mainly comes from primary research, timely literature reviews can lead to new synthetic insights and are often widely read [4] . For such summaries to be useful, however, they need to be compiled in a professional way [5] .
When starting from scratch, reviewing the literature can require a titanic amount of work. That is why researchers who have spent their career working on a certain research issue are in a perfect position to review that literature. Some graduate schools are now offering courses in reviewing the literature, given that most research students start their project by producing an overview of what has already been done on their research issue [6] . However, it is likely that most scientists have not thought in detail about how to approach and carry out a literature review.
Reviewing the literature requires the ability to juggle multiple tasks, from finding and evaluating relevant material to synthesising information from various sources, from critical thinking to paraphrasing, evaluating, and citation skills [7] . In this contribution, I share ten simple rules I learned working on about 25 literature reviews as a PhD and postdoctoral student. Ideas and insights also come from discussions with coauthors and colleagues, as well as feedback from reviewers and editors.
Rule 1: Define a Topic and Audience
How to choose which topic to review? There are so many issues in contemporary science that you could spend a lifetime of attending conferences and reading the literature just pondering what to review. On the one hand, if you take several years to choose, several other people may have had the same idea in the meantime. On the other hand, only a well-considered topic is likely to lead to a brilliant literature review [8] . The topic must at least be:
- interesting to you (ideally, you should have come across a series of recent papers related to your line of work that call for a critical summary),
- an important aspect of the field (so that many readers will be interested in the review and there will be enough material to write it), and
- a well-defined issue (otherwise you could potentially include thousands of publications, which would make the review unhelpful).
Ideas for potential reviews may come from papers providing lists of key research questions to be answered [9] , but also from serendipitous moments during desultory reading and discussions. In addition to choosing your topic, you should also select a target audience. In many cases, the topic (e.g., web services in computational biology) will automatically define an audience (e.g., computational biologists), but that same topic may also be of interest to neighbouring fields (e.g., computer science, biology, etc.).
Rule 2: Search and Re-search the Literature
After having chosen your topic and audience, start by checking the literature and downloading relevant papers. Five pieces of advice here:
- keep track of the search items you use (so that your search can be replicated [10] ),
- keep a list of papers whose pdfs you cannot access immediately (so as to retrieve them later with alternative strategies),
- use a paper management system (e.g., Mendeley, Papers, Qiqqa, Sente),
- define early in the process some criteria for exclusion of irrelevant papers (these criteria can then be described in the review to help define its scope), and
- do not just look for research papers in the area you wish to review, but also seek previous reviews.
The chances are high that someone will already have published a literature review ( Figure 1 ), if not exactly on the issue you are planning to tackle, at least on a related topic. If there are already a few or several reviews of the literature on your issue, my advice is not to give up, but to carry on with your own literature review,

The bottom-right situation (many literature reviews but few research papers) is not just a theoretical situation; it applies, for example, to the study of the impacts of climate change on plant diseases, where there appear to be more literature reviews than research studies [33] .
- discussing in your review the approaches, limitations, and conclusions of past reviews,
- trying to find a new angle that has not been covered adequately in the previous reviews, and
- incorporating new material that has inevitably accumulated since their appearance.
When searching the literature for pertinent papers and reviews, the usual rules apply:
- be thorough,
- use different keywords and database sources (e.g., DBLP, Google Scholar, ISI Proceedings, JSTOR Search, Medline, Scopus, Web of Science), and
- look at who has cited past relevant papers and book chapters.
Rule 3: Take Notes While Reading
If you read the papers first, and only afterwards start writing the review, you will need a very good memory to remember who wrote what, and what your impressions and associations were while reading each single paper. My advice is, while reading, to start writing down interesting pieces of information, insights about how to organize the review, and thoughts on what to write. This way, by the time you have read the literature you selected, you will already have a rough draft of the review.
Of course, this draft will still need much rewriting, restructuring, and rethinking to obtain a text with a coherent argument [11] , but you will have avoided the danger posed by staring at a blank document. Be careful when taking notes to use quotation marks if you are provisionally copying verbatim from the literature. It is advisable then to reformulate such quotes with your own words in the final draft. It is important to be careful in noting the references already at this stage, so as to avoid misattributions. Using referencing software from the very beginning of your endeavour will save you time.
Rule 4: Choose the Type of Review You Wish to Write
After having taken notes while reading the literature, you will have a rough idea of the amount of material available for the review. This is probably a good time to decide whether to go for a mini- or a full review. Some journals are now favouring the publication of rather short reviews focusing on the last few years, with a limit on the number of words and citations. A mini-review is not necessarily a minor review: it may well attract more attention from busy readers, although it will inevitably simplify some issues and leave out some relevant material due to space limitations. A full review will have the advantage of more freedom to cover in detail the complexities of a particular scientific development, but may then be left in the pile of the very important papers “to be read” by readers with little time to spare for major monographs.
There is probably a continuum between mini- and full reviews. The same point applies to the dichotomy of descriptive vs. integrative reviews. While descriptive reviews focus on the methodology, findings, and interpretation of each reviewed study, integrative reviews attempt to find common ideas and concepts from the reviewed material [12] . A similar distinction exists between narrative and systematic reviews: while narrative reviews are qualitative, systematic reviews attempt to test a hypothesis based on the published evidence, which is gathered using a predefined protocol to reduce bias [13] , [14] . When systematic reviews analyse quantitative results in a quantitative way, they become meta-analyses. The choice between different review types will have to be made on a case-by-case basis, depending not just on the nature of the material found and the preferences of the target journal(s), but also on the time available to write the review and the number of coauthors [15] .
Rule 5: Keep the Review Focused, but Make It of Broad Interest
Whether your plan is to write a mini- or a full review, it is good advice to keep it focused 16 , 17 . Including material just for the sake of it can easily lead to reviews that are trying to do too many things at once. The need to keep a review focused can be problematic for interdisciplinary reviews, where the aim is to bridge the gap between fields [18] . If you are writing a review on, for example, how epidemiological approaches are used in modelling the spread of ideas, you may be inclined to include material from both parent fields, epidemiology and the study of cultural diffusion. This may be necessary to some extent, but in this case a focused review would only deal in detail with those studies at the interface between epidemiology and the spread of ideas.
While focus is an important feature of a successful review, this requirement has to be balanced with the need to make the review relevant to a broad audience. This square may be circled by discussing the wider implications of the reviewed topic for other disciplines.
Rule 6: Be Critical and Consistent
Reviewing the literature is not stamp collecting. A good review does not just summarize the literature, but discusses it critically, identifies methodological problems, and points out research gaps [19] . After having read a review of the literature, a reader should have a rough idea of:
- the major achievements in the reviewed field,
- the main areas of debate, and
- the outstanding research questions.
It is challenging to achieve a successful review on all these fronts. A solution can be to involve a set of complementary coauthors: some people are excellent at mapping what has been achieved, some others are very good at identifying dark clouds on the horizon, and some have instead a knack at predicting where solutions are going to come from. If your journal club has exactly this sort of team, then you should definitely write a review of the literature! In addition to critical thinking, a literature review needs consistency, for example in the choice of passive vs. active voice and present vs. past tense.
Rule 7: Find a Logical Structure
Like a well-baked cake, a good review has a number of telling features: it is worth the reader's time, timely, systematic, well written, focused, and critical. It also needs a good structure. With reviews, the usual subdivision of research papers into introduction, methods, results, and discussion does not work or is rarely used. However, a general introduction of the context and, toward the end, a recapitulation of the main points covered and take-home messages make sense also in the case of reviews. For systematic reviews, there is a trend towards including information about how the literature was searched (database, keywords, time limits) [20] .
How can you organize the flow of the main body of the review so that the reader will be drawn into and guided through it? It is generally helpful to draw a conceptual scheme of the review, e.g., with mind-mapping techniques. Such diagrams can help recognize a logical way to order and link the various sections of a review [21] . This is the case not just at the writing stage, but also for readers if the diagram is included in the review as a figure. A careful selection of diagrams and figures relevant to the reviewed topic can be very helpful to structure the text too [22] .
Rule 8: Make Use of Feedback
Reviews of the literature are normally peer-reviewed in the same way as research papers, and rightly so [23] . As a rule, incorporating feedback from reviewers greatly helps improve a review draft. Having read the review with a fresh mind, reviewers may spot inaccuracies, inconsistencies, and ambiguities that had not been noticed by the writers due to rereading the typescript too many times. It is however advisable to reread the draft one more time before submission, as a last-minute correction of typos, leaps, and muddled sentences may enable the reviewers to focus on providing advice on the content rather than the form.
Feedback is vital to writing a good review, and should be sought from a variety of colleagues, so as to obtain a diversity of views on the draft. This may lead in some cases to conflicting views on the merits of the paper, and on how to improve it, but such a situation is better than the absence of feedback. A diversity of feedback perspectives on a literature review can help identify where the consensus view stands in the landscape of the current scientific understanding of an issue [24] .
Rule 9: Include Your Own Relevant Research, but Be Objective
In many cases, reviewers of the literature will have published studies relevant to the review they are writing. This could create a conflict of interest: how can reviewers report objectively on their own work [25] ? Some scientists may be overly enthusiastic about what they have published, and thus risk giving too much importance to their own findings in the review. However, bias could also occur in the other direction: some scientists may be unduly dismissive of their own achievements, so that they will tend to downplay their contribution (if any) to a field when reviewing it.
In general, a review of the literature should neither be a public relations brochure nor an exercise in competitive self-denial. If a reviewer is up to the job of producing a well-organized and methodical review, which flows well and provides a service to the readership, then it should be possible to be objective in reviewing one's own relevant findings. In reviews written by multiple authors, this may be achieved by assigning the review of the results of a coauthor to different coauthors.
Rule 10: Be Up-to-Date, but Do Not Forget Older Studies
Given the progressive acceleration in the publication of scientific papers, today's reviews of the literature need awareness not just of the overall direction and achievements of a field of inquiry, but also of the latest studies, so as not to become out-of-date before they have been published. Ideally, a literature review should not identify as a major research gap an issue that has just been addressed in a series of papers in press (the same applies, of course, to older, overlooked studies (“sleeping beauties” [26] )). This implies that literature reviewers would do well to keep an eye on electronic lists of papers in press, given that it can take months before these appear in scientific databases. Some reviews declare that they have scanned the literature up to a certain point in time, but given that peer review can be a rather lengthy process, a full search for newly appeared literature at the revision stage may be worthwhile. Assessing the contribution of papers that have just appeared is particularly challenging, because there is little perspective with which to gauge their significance and impact on further research and society.
Inevitably, new papers on the reviewed topic (including independently written literature reviews) will appear from all quarters after the review has been published, so that there may soon be the need for an updated review. But this is the nature of science [27] – [32] . I wish everybody good luck with writing a review of the literature.
Acknowledgments
Many thanks to M. Barbosa, K. Dehnen-Schmutz, T. Döring, D. Fontaneto, M. Garbelotto, O. Holdenrieder, M. Jeger, D. Lonsdale, A. MacLeod, P. Mills, M. Moslonka-Lefebvre, G. Stancanelli, P. Weisberg, and X. Xu for insights and discussions, and to P. Bourne, T. Matoni, and D. Smith for helpful comments on a previous draft.
Funding Statement
This work was funded by the French Foundation for Research on Biodiversity (FRB) through its Centre for Synthesis and Analysis of Biodiversity data (CESAB), as part of the NETSEED research project. The funders had no role in the preparation of the manuscript.
- Privacy Policy
Home » Literature Review – Types Writing Guide and Examples
Literature Review – Types Writing Guide and Examples
Table of Contents

Literature Review
Definition:
A literature review is a comprehensive and critical analysis of the existing literature on a particular topic or research question. It involves identifying, evaluating, and synthesizing relevant literature, including scholarly articles, books, and other sources, to provide a summary and critical assessment of what is known about the topic.
Types of Literature Review
Types of Literature Review are as follows:
- Narrative literature review : This type of review involves a comprehensive summary and critical analysis of the available literature on a particular topic or research question. It is often used as an introductory section of a research paper.
- Systematic literature review: This is a rigorous and structured review that follows a pre-defined protocol to identify, evaluate, and synthesize all relevant studies on a specific research question. It is often used in evidence-based practice and systematic reviews.
- Meta-analysis: This is a quantitative review that uses statistical methods to combine data from multiple studies to derive a summary effect size. It provides a more precise estimate of the overall effect than any individual study.
- Scoping review: This is a preliminary review that aims to map the existing literature on a broad topic area to identify research gaps and areas for further investigation.
- Critical literature review : This type of review evaluates the strengths and weaknesses of the existing literature on a particular topic or research question. It aims to provide a critical analysis of the literature and identify areas where further research is needed.
- Conceptual literature review: This review synthesizes and integrates theories and concepts from multiple sources to provide a new perspective on a particular topic. It aims to provide a theoretical framework for understanding a particular research question.
- Rapid literature review: This is a quick review that provides a snapshot of the current state of knowledge on a specific research question or topic. It is often used when time and resources are limited.
- Thematic literature review : This review identifies and analyzes common themes and patterns across a body of literature on a particular topic. It aims to provide a comprehensive overview of the literature and identify key themes and concepts.
- Realist literature review: This review is often used in social science research and aims to identify how and why certain interventions work in certain contexts. It takes into account the context and complexities of real-world situations.
- State-of-the-art literature review : This type of review provides an overview of the current state of knowledge in a particular field, highlighting the most recent and relevant research. It is often used in fields where knowledge is rapidly evolving, such as technology or medicine.
- Integrative literature review: This type of review synthesizes and integrates findings from multiple studies on a particular topic to identify patterns, themes, and gaps in the literature. It aims to provide a comprehensive understanding of the current state of knowledge on a particular topic.
- Umbrella literature review : This review is used to provide a broad overview of a large and diverse body of literature on a particular topic. It aims to identify common themes and patterns across different areas of research.
- Historical literature review: This type of review examines the historical development of research on a particular topic or research question. It aims to provide a historical context for understanding the current state of knowledge on a particular topic.
- Problem-oriented literature review : This review focuses on a specific problem or issue and examines the literature to identify potential solutions or interventions. It aims to provide practical recommendations for addressing a particular problem or issue.
- Mixed-methods literature review : This type of review combines quantitative and qualitative methods to synthesize and analyze the available literature on a particular topic. It aims to provide a more comprehensive understanding of the research question by combining different types of evidence.
Parts of Literature Review
Parts of a literature review are as follows:
Introduction
The introduction of a literature review typically provides background information on the research topic and why it is important. It outlines the objectives of the review, the research question or hypothesis, and the scope of the review.
Literature Search
This section outlines the search strategy and databases used to identify relevant literature. The search terms used, inclusion and exclusion criteria, and any limitations of the search are described.
Literature Analysis
The literature analysis is the main body of the literature review. This section summarizes and synthesizes the literature that is relevant to the research question or hypothesis. The review should be organized thematically, chronologically, or by methodology, depending on the research objectives.
Critical Evaluation
Critical evaluation involves assessing the quality and validity of the literature. This includes evaluating the reliability and validity of the studies reviewed, the methodology used, and the strength of the evidence.
The conclusion of the literature review should summarize the main findings, identify any gaps in the literature, and suggest areas for future research. It should also reiterate the importance of the research question or hypothesis and the contribution of the literature review to the overall research project.
The references list includes all the sources cited in the literature review, and follows a specific referencing style (e.g., APA, MLA, Harvard).
How to write Literature Review
Here are some steps to follow when writing a literature review:
- Define your research question or topic : Before starting your literature review, it is essential to define your research question or topic. This will help you identify relevant literature and determine the scope of your review.
- Conduct a comprehensive search: Use databases and search engines to find relevant literature. Look for peer-reviewed articles, books, and other academic sources that are relevant to your research question or topic.
- Evaluate the sources: Once you have found potential sources, evaluate them critically to determine their relevance, credibility, and quality. Look for recent publications, reputable authors, and reliable sources of data and evidence.
- Organize your sources: Group the sources by theme, method, or research question. This will help you identify similarities and differences among the literature, and provide a structure for your literature review.
- Analyze and synthesize the literature : Analyze each source in depth, identifying the key findings, methodologies, and conclusions. Then, synthesize the information from the sources, identifying patterns and themes in the literature.
- Write the literature review : Start with an introduction that provides an overview of the topic and the purpose of the literature review. Then, organize the literature according to your chosen structure, and analyze and synthesize the sources. Finally, provide a conclusion that summarizes the key findings of the literature review, identifies gaps in knowledge, and suggests areas for future research.
- Edit and proofread: Once you have written your literature review, edit and proofread it carefully to ensure that it is well-organized, clear, and concise.
Examples of Literature Review
Here’s an example of how a literature review can be conducted for a thesis on the topic of “ The Impact of Social Media on Teenagers’ Mental Health”:
- Start by identifying the key terms related to your research topic. In this case, the key terms are “social media,” “teenagers,” and “mental health.”
- Use academic databases like Google Scholar, JSTOR, or PubMed to search for relevant articles, books, and other publications. Use these keywords in your search to narrow down your results.
- Evaluate the sources you find to determine if they are relevant to your research question. You may want to consider the publication date, author’s credentials, and the journal or book publisher.
- Begin reading and taking notes on each source, paying attention to key findings, methodologies used, and any gaps in the research.
- Organize your findings into themes or categories. For example, you might categorize your sources into those that examine the impact of social media on self-esteem, those that explore the effects of cyberbullying, and those that investigate the relationship between social media use and depression.
- Synthesize your findings by summarizing the key themes and highlighting any gaps or inconsistencies in the research. Identify areas where further research is needed.
- Use your literature review to inform your research questions and hypotheses for your thesis.
For example, after conducting a literature review on the impact of social media on teenagers’ mental health, a thesis might look like this:
“Using a mixed-methods approach, this study aims to investigate the relationship between social media use and mental health outcomes in teenagers. Specifically, the study will examine the effects of cyberbullying, social comparison, and excessive social media use on self-esteem, anxiety, and depression. Through an analysis of survey data and qualitative interviews with teenagers, the study will provide insight into the complex relationship between social media use and mental health outcomes, and identify strategies for promoting positive mental health outcomes in young people.”
Reference: Smith, J., Jones, M., & Lee, S. (2019). The effects of social media use on adolescent mental health: A systematic review. Journal of Adolescent Health, 65(2), 154-165. doi:10.1016/j.jadohealth.2019.03.024
Reference Example: Author, A. A., Author, B. B., & Author, C. C. (Year). Title of article. Title of Journal, volume number(issue number), page range. doi:0000000/000000000000 or URL
Applications of Literature Review
some applications of literature review in different fields:
- Social Sciences: In social sciences, literature reviews are used to identify gaps in existing research, to develop research questions, and to provide a theoretical framework for research. Literature reviews are commonly used in fields such as sociology, psychology, anthropology, and political science.
- Natural Sciences: In natural sciences, literature reviews are used to summarize and evaluate the current state of knowledge in a particular field or subfield. Literature reviews can help researchers identify areas where more research is needed and provide insights into the latest developments in a particular field. Fields such as biology, chemistry, and physics commonly use literature reviews.
- Health Sciences: In health sciences, literature reviews are used to evaluate the effectiveness of treatments, identify best practices, and determine areas where more research is needed. Literature reviews are commonly used in fields such as medicine, nursing, and public health.
- Humanities: In humanities, literature reviews are used to identify gaps in existing knowledge, develop new interpretations of texts or cultural artifacts, and provide a theoretical framework for research. Literature reviews are commonly used in fields such as history, literary studies, and philosophy.
Role of Literature Review in Research
Here are some applications of literature review in research:
- Identifying Research Gaps : Literature review helps researchers identify gaps in existing research and literature related to their research question. This allows them to develop new research questions and hypotheses to fill those gaps.
- Developing Theoretical Framework: Literature review helps researchers develop a theoretical framework for their research. By analyzing and synthesizing existing literature, researchers can identify the key concepts, theories, and models that are relevant to their research.
- Selecting Research Methods : Literature review helps researchers select appropriate research methods and techniques based on previous research. It also helps researchers to identify potential biases or limitations of certain methods and techniques.
- Data Collection and Analysis: Literature review helps researchers in data collection and analysis by providing a foundation for the development of data collection instruments and methods. It also helps researchers to identify relevant data sources and identify potential data analysis techniques.
- Communicating Results: Literature review helps researchers to communicate their results effectively by providing a context for their research. It also helps to justify the significance of their findings in relation to existing research and literature.
Purpose of Literature Review
Some of the specific purposes of a literature review are as follows:
- To provide context: A literature review helps to provide context for your research by situating it within the broader body of literature on the topic.
- To identify gaps and inconsistencies: A literature review helps to identify areas where further research is needed or where there are inconsistencies in the existing literature.
- To synthesize information: A literature review helps to synthesize the information from multiple sources and present a coherent and comprehensive picture of the current state of knowledge on the topic.
- To identify key concepts and theories : A literature review helps to identify key concepts and theories that are relevant to your research question and provide a theoretical framework for your study.
- To inform research design: A literature review can inform the design of your research study by identifying appropriate research methods, data sources, and research questions.
Characteristics of Literature Review
Some Characteristics of Literature Review are as follows:
- Identifying gaps in knowledge: A literature review helps to identify gaps in the existing knowledge and research on a specific topic or research question. By analyzing and synthesizing the literature, you can identify areas where further research is needed and where new insights can be gained.
- Establishing the significance of your research: A literature review helps to establish the significance of your own research by placing it in the context of existing research. By demonstrating the relevance of your research to the existing literature, you can establish its importance and value.
- Informing research design and methodology : A literature review helps to inform research design and methodology by identifying the most appropriate research methods, techniques, and instruments. By reviewing the literature, you can identify the strengths and limitations of different research methods and techniques, and select the most appropriate ones for your own research.
- Supporting arguments and claims: A literature review provides evidence to support arguments and claims made in academic writing. By citing and analyzing the literature, you can provide a solid foundation for your own arguments and claims.
- I dentifying potential collaborators and mentors: A literature review can help identify potential collaborators and mentors by identifying researchers and practitioners who are working on related topics or using similar methods. By building relationships with these individuals, you can gain valuable insights and support for your own research and practice.
- Keeping up-to-date with the latest research : A literature review helps to keep you up-to-date with the latest research on a specific topic or research question. By regularly reviewing the literature, you can stay informed about the latest findings and developments in your field.
Advantages of Literature Review
There are several advantages to conducting a literature review as part of a research project, including:
- Establishing the significance of the research : A literature review helps to establish the significance of the research by demonstrating the gap or problem in the existing literature that the study aims to address.
- Identifying key concepts and theories: A literature review can help to identify key concepts and theories that are relevant to the research question, and provide a theoretical framework for the study.
- Supporting the research methodology : A literature review can inform the research methodology by identifying appropriate research methods, data sources, and research questions.
- Providing a comprehensive overview of the literature : A literature review provides a comprehensive overview of the current state of knowledge on a topic, allowing the researcher to identify key themes, debates, and areas of agreement or disagreement.
- Identifying potential research questions: A literature review can help to identify potential research questions and areas for further investigation.
- Avoiding duplication of research: A literature review can help to avoid duplication of research by identifying what has already been done on a topic, and what remains to be done.
- Enhancing the credibility of the research : A literature review helps to enhance the credibility of the research by demonstrating the researcher’s knowledge of the existing literature and their ability to situate their research within a broader context.
Limitations of Literature Review
Limitations of Literature Review are as follows:
- Limited scope : Literature reviews can only cover the existing literature on a particular topic, which may be limited in scope or depth.
- Publication bias : Literature reviews may be influenced by publication bias, which occurs when researchers are more likely to publish positive results than negative ones. This can lead to an incomplete or biased picture of the literature.
- Quality of sources : The quality of the literature reviewed can vary widely, and not all sources may be reliable or valid.
- Time-limited: Literature reviews can become quickly outdated as new research is published, making it difficult to keep up with the latest developments in a field.
- Subjective interpretation : Literature reviews can be subjective, and the interpretation of the findings can vary depending on the researcher’s perspective or bias.
- Lack of original data : Literature reviews do not generate new data, but rather rely on the analysis of existing studies.
- Risk of plagiarism: It is important to ensure that literature reviews do not inadvertently contain plagiarism, which can occur when researchers use the work of others without proper attribution.
About the author

Muhammad Hassan
Researcher, Academic Writer, Web developer
You may also like

Research Recommendations – Examples and Writing...

Critical Analysis – Types, Examples and Writing...

Table of Contents – Types, Formats, Examples

Dissertation vs Thesis – Key Differences

Survey Instruments – List and Their Uses

Scope of the Research – Writing Guide and...
Purdue Online Writing Lab Purdue OWL® College of Liberal Arts
Literature Topics and Research

Welcome to the Purdue OWL
This page is brought to you by the OWL at Purdue University. When printing this page, you must include the entire legal notice.
Copyright ©1995-2018 by The Writing Lab & The OWL at Purdue and Purdue University. All rights reserved. This material may not be published, reproduced, broadcast, rewritten, or redistributed without permission. Use of this site constitutes acceptance of our terms and conditions of fair use.
What kinds of topics are good ones?
The best topics are ones that originate out of your own reading of a work of literature, but here are some common approaches to consider:
- A discussion of a work's characters: are they realistic, symbolic, historically-based?
- A comparison/contrast of the choices different authors or characters make in a work
- A reading of a work based on an outside philosophical perspective (Ex. how would a Freudian read Hamlet ?)
- A study of the sources or historical events that occasioned a particular work (Ex. comparing G.B. Shaw's Pygmalion with the original Greek myth of Pygmalion)
- An analysis of a specific image occurring in several works (Ex. the use of moon imagery in certain plays, poems, novels)
- A "deconstruction" of a particular work (Ex. unfolding an underlying racist worldview in Joseph Conrad's Heart of Darkness )
- A reading from a political perspective (Ex. how would a Marxist read William Blake's "London"?)
- A study of the social, political, or economic context in which a work was written — how does the context influence the work?
How do I start research?
Once you have decided on an interesting topic and work (or works), the best place to start is probably the Internet. Here you can usually find basic biographical data on authors, brief summaries of works, possibly some rudimentary analyses, and even bibliographies of sources related to your topic.
The Internet, however, rarely offers serious direct scholarship; you will have to use sources found in the library, sources like journal articles and scholarly books, to get information that you can use to build your own scholarship-your literary paper. Consult the library's on-line catalog and the MLA Periodical Index. Avoid citing dictionary or encyclopedic sources in your final paper.
How do I use the information I find?
The secondary sources you find are only to be used as an aid. Your thoughts should make up most of the essay. As you develop your thesis, you will bring in the ideas of the scholars to back up what you have already said.
For example, say you are arguing that Huck Finn is a Christ figure ; that's your basic thesis. You give evidence from the novel that allows this reading, and then, at the right place, you might say the following, a paraphrase:
According to Susan Thomas, Huck sacrifices himself because he wants to set Jim free (129).
If the scholar states an important idea in a memorable way, use a direct quote.
"Huck's altruism and feelings of compassion for Jim force him to surrender to the danger" (Thomas 129).
Either way, you will then link that idea to your thesis.
Something went wrong when searching for seed articles. Please try again soon.
No articles were found for that search term.
Author, year The title of the article goes here
LITERATURE REVIEW SOFTWARE FOR BETTER RESEARCH
“Litmaps is a game changer for finding novel literature... it has been invaluable for my productivity.... I also got my PhD student to use it and they also found it invaluable, finding several gaps they missed”
Varun Venkatesh
Austin Health, Australia

As a full-time researcher, Litmaps has become an indispensable tool in my arsenal. The Seed Maps and Discover features of Litmaps have transformed my literature review process, streamlining the identification of key citations while revealing previously overlooked relevant literature, ensuring no crucial connection goes unnoticed. A true game-changer indeed!
Ritwik Pandey
Doctoral Research Scholar – Sri Sathya Sai Institute of Higher Learning

Using Litmaps for my research papers has significantly improved my workflow. Typically, I start with a single paper related to my topic. Whenever I find an interesting work, I add it to my search. From there, I can quickly cover my entire Related Work section.
David Fischer
Research Associate – University of Applied Sciences Kempten
“It's nice to get a quick overview of related literature. Really easy to use, and it helps getting on top of the often complicated structures of referencing”
Christoph Ludwig
Technische Universität Dresden, Germany
“This has helped me so much in researching the literature. Currently, I am beginning to investigate new fields and this has helped me hugely”
Aran Warren
Canterbury University, NZ
“I can’t live without you anymore! I also recommend you to my students.”
Professor at The Chinese University of Hong Kong
“Seeing my literature list as a network enhances my thinking process!”
Katholieke Universiteit Leuven, Belgium
“Incredibly useful tool to get to know more literature, and to gain insight in existing research”
KU Leuven, Belgium
“As a student just venturing into the world of lit reviews, this is a tool that is outstanding and helping me find deeper results for my work.”
Franklin Jeffers
South Oregon University, USA
“Any researcher could use it! The paper recommendations are great for anyone and everyone”
Swansea University, Wales
“This tool really helped me to create good bibtex references for my research papers”
Ali Mohammed-Djafari
Director of Research at LSS-CNRS, France
“Litmaps is extremely helpful with my research. It helps me organize each one of my projects and see how they relate to each other, as well as to keep up to date on publications done in my field”
Daniel Fuller
Clarkson University, USA
As a person who is an early researcher and identifies as dyslexic, I can say that having research articles laid out in the date vs cite graph format is much more approachable than looking at a standard database interface. I feel that the maps Litmaps offers lower the barrier of entry for researchers by giving them the connections between articles spaced out visually. This helps me orientate where a paper is in the history of a field. Thus, new researchers can look at one of Litmap's "seed maps" and have the same information as hours of digging through a database.
Baylor Fain
Postdoctoral Associate – University of Florida

We use cookies to ensure that we give you the best experience on our website. If you continue to use this site we will assume that you are happy with it.

Writing the Literature Review
Barry Mauer and John Venecek
- The Literature Review
What is the Purpose of the Review?
What is the scope of the review, strategies for getting started, types of literature reviews, composition guidelines, how to locate reviews by discipline, key takeaways.
We also provide the following activities:
Types of Literature Reviews [Refresher]
Writing the literature review [refresher], the literature review [1].
Conducting a literary studies research project involves time and effort, with much of it going towards the development of a literature review . A literature review might fill several pages of your research paper and usually appears soon after an introduction but before you present your detailed argument. A literature review provides your audience with an overview of the available research about your area(s) of study, including the literary work, your theory, and methodology. The literature review demonstrates how these scholarly discussions have changed over time, and it allows you to position your research in relation to research that has come before yours. Your aim is to present the discussion up to this point. Depending on the nature of the assignment, you may also include your critical commentary on prior research, noting among this material the weaker and stronger arguments, breakthroughs and dead ends, blind spots and opportunities, the invention of key terms and methods, mistakes as well as misreadings, and so on.
Once you have gathered the research materials you need for your literature review, you have another task: conducting an analysis on the research to see where your original contribution fits into the scholarly conversation. As the saying goes, “we are standing on the shoulders of giants.” Your job is to show a portrait of these giants to your audience, and to show how your work relates to the portrait. On many scholarly topics, literature reviews already exist. You may refer to such existing reviews within your own, indicating any materials might have been overlooked, new developments that have arisen since the publication of the existing literature review, and new perspectives or insights you have about the materials.
Some beginning researchers try to tear down the work of other researchers in an effort to make their own work look good by comparison. It rarely works. First, it tends to make your audience skeptical of your claims. Second, it ignores the fact that even the mistakes, blind spots, and failures of other researchers contribute something to our knowledge. Albert Einstein didn’t disrespect Sir Isaac Newton by saying Newton’s theory of space was wrong and terrible and that Einstein’s own theory was great by comparison. He built upon Newton’s work, showing how it could be improved. If, however, a researcher willfully set out to deceive or distort or to tear down the work of other scholars without good reason, then their work does not deserve such deference.
Most literature reviews appear after the introduction. It presents your reader with relevant information about the scholarly discussion up to now. Later in your paper, you discuss your contribution. Before you begin work on your literature review, let’s discuss what we mean by “literature”; understand the purpose and scope of the review; establish criteria for selecting, organizing, and interpreting your findings; and discuss how to connect your findings to your research question.
Many students seek to “find sources that agree with my claim or idea.” That approach is too narrow, in our view. If we use such an approach, we may get the following results:
- Because we can find sources that agree with almost any claim, readers will wonder whether your claims are weak and the sources are cherry picked.
- While literary scholars sometimes cite authorities to support their claims, they don’t rely only on authority. They respect authority, but not too much. Your own claims need to rely more on evidence (from the literary text, historical and biographical information), and your critical and creative reasoning skills.
- Scholarship is a conversation; thus, the goal is less about finding agreement and more about joining the conversation with the aim of making a valuable contribution to the discussion.
The literature review provides your reader with an overview of the existing research about your topic or problem. It provides the context necessary for your reader to catch up with the scholarly conversation and then to appreciate the value of your contribution to it. The literature review sharpens the focus of your research and demonstrates your knowledge and understanding of the scholarly conversation around your topic, which, in turn, helps establish your credibility as a researcher.
Creating the literature review involves more than gathering citations. It is a qualitative process through which you will discover what is already known about your topic, and identify the key authorities, methods, and theoretical foundations, so you can begin to position your contributions within the scholarly conversation.
Defining the scope of your review will also help you establish criteria to determine the relevance of the sources you are finding. At this stage, you are not reading in-depth; instead, you are skimming through what has already been published and identifying the major concepts, theories, methodologies, and methods present within these published works. You should also be identifying connections, tensions, and contradictions within the already published works of your topic or problem. This involves building on the knowledge of others and understanding what methods, measures, and models we have inherited from previous researchers in our field.
Literature Reviews: Common Errors Made When Conducting a Literature Review [12 min 22 sec]
Video provided courtesy of the Center for Quality Research (CQR)
A literature review helps your reader understand the relationship of your research project to the work of other scholars. It covers the existing knowledge about a problem, and allows you to show the relevance/significance of your contribution to the discussion. Your reader may or may not have read scholarly literature about the theories, methodologies, and literary works you are discussing. But they want to know that you have read it and have thought about it. Your literature review provides not only a summary of the existing scholarship for readers; it also offers your perspective on it.
Begin your work on the literature review by synthesizing the various sources in your annotated bibliography .
For advice on Synthesizing Sources, consider the following from The Purdue Online Writing Lab: [2]
Note that synthesizing is not the same as summarizing .
- A summary restates the information in one or more sources without providing new insight or reaching new conclusions.
- A synthesis draws on multiple sources to reach a broader conclusion.
- Don’t force a relationship between sources if there isn’t one. Not all of your sources have to complement one another.
- Do your best to highlight the relationships between sources in very clear ways.
- Don’t ignore any outliers in your research. It’s important to take note of every perspective (even those that disagree with your broader conclusions).
Not all humanities research projects contain literature reviews, but many do. Keep in mind that the type of literature review you choose (see list below) pertains to the secondary research – other scholarly sources – and not to the primary literary work. For instance, a literature review about Kate Chopin’s writing will be your thoughts about the scholarship on Chopin and not about Chopin’s text itself. You are summarizing what you see in the scholarly literature about Chopin’s writing. The literature review puts you in the position of authority not just on Chopin’s writing but on the scholarship about her writing. You are seeking to understand what scholars have said about her work. Scholars might belong to different schools of thought (psychoanalytic, feminist, Marxist, etc.). They might make different arguments about Chopin. They might use different methodological approaches.
If your research involves two or more theories, such as psychology and genre studies, you may need to create multiple literature reviews, one for each theory or methodology. If the theories overlap with each other significantly (i.e., Marxism and Cultural Studies), you may combine them. Your literature review need not include everything about the subject area – you would need to write a book to cover a single theory – but only those concepts and methods that are most relevant to your research problem.
Factors to Consider When Developing Your Literature Review
- Determine the Scope : How broad or narrow should your literature review be? You may want to focus on recent scholarship only, or on a particular school of thought in the literature. Your scope is determined by your purpose; what is it you aim to achieve with your research?
- Establish Criteria : We discussed the importance of defining the purpose and scope of your review on the previous page, but it’s worth reviewing here as well. This step will help you establish important criteria and focus your searching. For example, how many sources will you need? What types of sources (primary, secondary, statistics, media)? Is currency important? Do you know who the prominent authors or theorists are in your subject area? Take some time to map out these or other important factors before you begin searching journals and databases.
- Consider Your Audience : Unlike a work cited page or an annotated bibliography, both of which are lists of sources, a literature review is essayistic and can be considered a precursor to your final paper. Therefore, it should be written in your own voice, and it should be geared toward a specific audience. Considering audience during this early stage will help focus your final paper as well.
- Find Models : We’ll discuss the different types of literature reviews and how to locate examples in the section below. However, even if you’re undecided about what type of review will work best for you, you may want to review some example literature reviews to get a sense of what they look like before you begin your own.
One piece of advice before starting: look for existing literature reviews on your area of scholarship. You can build on the work that other scholars have put into reviewing the scholarly literature. There’s no need to completely “reinvent the wheel” if some of the work is already done.
Scholars sometimes publish “stand-alone” literature reviews that are not part of a larger work; such literature reviews are valuable contributions to the field, as they summarize the state of knowledge for other scholars.
Maria J. Grant and Andrew Booth’s “A Typology of Reviews” identifies 14 distinct types of literature reviews. Further, the UCLA library created a chart to complement the article and for easy comparison of those 14 types of reviews. This section provides a brief summary of the most common literature reviews. For a more complete analysis, please see the full article and the chart .
To choose the most appropriate structure, put yourself in your reader’s shoes and think through their need for information. The literature review is about providing context for your contribution. How much context do people need? Keep it to the minimum necessary; compressing a lot of information into a small amount of text is a must.
These structures are not meant to be straightjackets but tools to help you organize your research. If you find that the tool is working, then keep using it. If not, switch tools or modify the one you are using. Keep in mind that the types of literature reviews are just different ways of organizing information. So, you can discuss literary trends without organizing your review of secondary literature by trend; your discussion can be organized by theory or theme, for examples. In our literature reviews, we are not recounting other scholars’ arguments at length but merely providing key concepts so we can summarize the discussion so far and position our own claims. You don’t have to adhere strictly to one structure or another. They are just organizing tools that help you manage your material (and help your reader make sense of it).
Types of Reviews
- Traditional or narrative reviews : This approach will generate a comprehensive, critical analysis of the published research on your topic. However, rather than merely compiling as many sources as possible, use this approach to establish a theoretical framework for your paper, establish trends, and identify gaps in the research. This process should bring your research question into clearer focus and help define a thesis that you will argue for in your paper. This is perhaps the most common and general type of literature review. The examples listed below are all designed to serve a more specific purpose.
- Argumentative : The purpose of an argumentative literature review is to select sources for the purpose of supporting or refuting a specific claim. While this type of review can help the author make a strong case for or against an issue, they can also be prone to claims of bias. Later in this textbook, we will read about the distinction between warranted and unwarranted bias . One is ok and the other is not.
- Chronological : A chronological review is used when the author wants to demonstrate the progression of how a theory, methodology, or issue has progressed over time. This method is most effective when there is a clear chronological path to the research about a specific historical event or trend as opposed to a more recursive theoretical concept.
- By trend : This is similar to the chronological approach except it focuses on clearly-defined trends rather than date ranges. This would be most appropriate if you want to illustrate changing perspectives or attitudes about a given issue when specific date ranges are less important than the ebb and flow of the trend.
- Thematic : In this type of literature review, the author will select specific themes that he or she feels are important to understanding a larger topic or concept. Then, the author will organize the sources around those themes, which are often based on relevance or importance. The value of this method is that the process of organizing the review by theme is similar to constructing an argument. This can help the author see how resources connect to each other and determine how as well as why specific sources support their thesis.
- Theoretical : The goal of this type of review is to examine how theory has shaped the research on a given topic. It establishes existing theoretical models, their connections, and how extensively they have been developed in the published research. For example, Jada applied critical race theory to her analysis of Sonny’s Blues , but she might also consider conducting a more comprehensive review of other theoretical frameworks such as feminism, Marxism, or postmodernism. Doing so could provide insight into alternate readings, and help her identify theoretical gaps such as unexplored or under-developed approaches to Baldwin’s work.
- Methodological : The approach focuses on the various methodologies used by researchers in a specific area rather than an analysis of their findings. In this case, you would create a framework of approaches to data collection related to your topic or research question. This is perhaps more common in education or the social and hard sciences where published research often includes a methods section, but it is sometimes appropriate for the digital humanities as well.
- Scoping : The aim of a scoping review is to provide a comprehensive overview or map of the published research or evidence related to a research question. This might be considered a prelude to a systematic review that would take the scoping review one step further toward answering a clearly defined research question. See below for more details.
- Systematic : The systematic review is most appropriate when you have a clearly-defined research question and have established criteria for the types of sources you need. In this way, the systematic review is less exploratory than other types of reviews. Rather, it is comprehensive, strategic, and focused on answering a specific research question. For this reason, the systematic review is more common in the health and social sciences, where comprehensiveness is more important. Literature reviews in the Humanities are not usually exhaustive but tend to show only the most representative or salient developments in the scholarship.
- Meta-analysis : Does your research deal with statistics or large amounts of data? If so, then a meta-analysis might be best for you rather than providing a critical review, the meta-analysis will summarize and synthesize the results of numerous studies that involve statistics or data to provide a more comprehensive picture than would be possible from just one study.
An argumentative literature review presents and takes sides in scholarly arguments about the literary work. It makes arguments about other scholars’ work. It does not necessarily involve a claim that the literary work is itself making an argument. Likewise, a chronological literature review presents the scholarly literature in chronological order.
You don’t need to keep strictly to one type. Scholars often combine features from various types of literature reviews. A sample review that combines the follow types –
- Argumentative
- Theoretical
- Methodological
– is the excellent work of Eiranen, Reetta, Mari Hatavara, Ville Kivimäki, Maria Mäkelä & Raisa Maria Toivo (2022) “ Narrative and Experience: Interdisciplinary Methodologies between History and Narratology , ” Scandinavian Journal of History , 47:1, 1-15
When writing your literature review, please follow these pointers:
- Conduct systematic searches
- Use Evidence
- Be Selective
- Use Quotes Sparingly
- Summarize & Synthesize
- Use Caution when Paraphrasing
- Use Your Own Voice
Advice from James Mason University’s “Literature Reviews: An Overview”
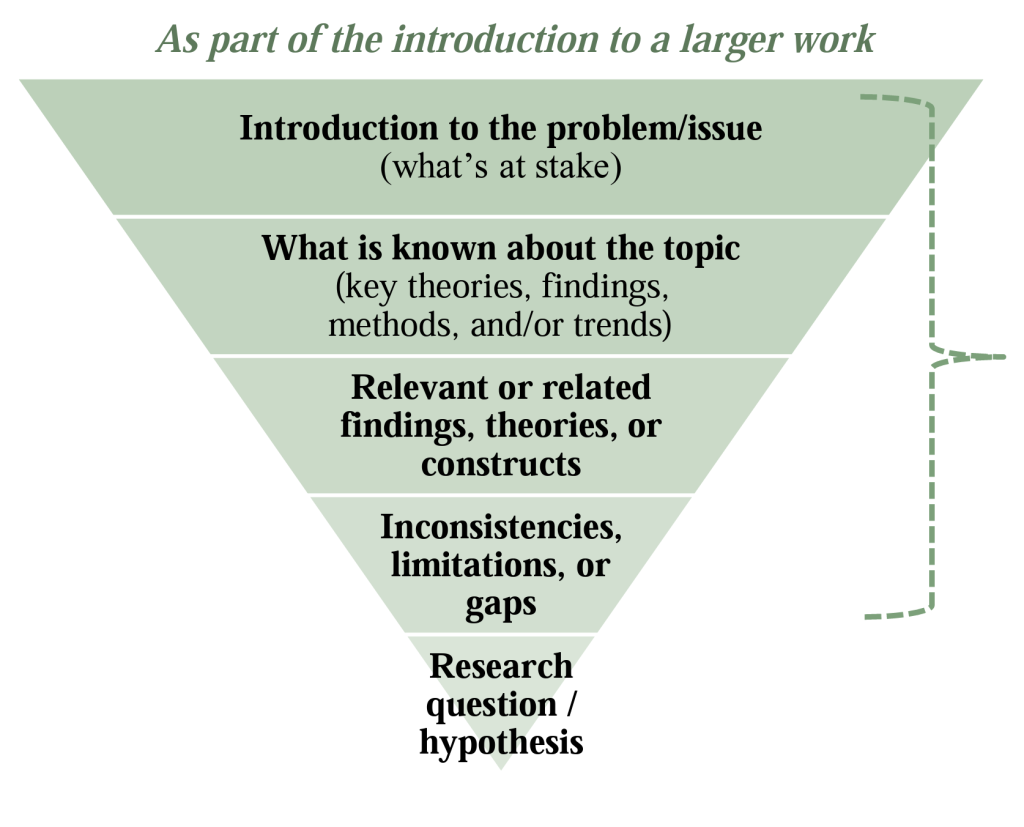
A note on synthesizing : Don’t make the common mistake of summarizing individual studies or articles one after the other. The goal is to synthesize — that is, to make observations about groups of studies. Synthesis often uses language like this:
- Much of the literature on [topic x ] focuses on [major themes].
- In recent years, researchers have begun investigating [facets a , b , and c ] of [topic x ].
- The studies in this review of [topic x ] confirm / suggest / call into question / support [idea / practice / finding / method / theory / guideline y ].
- In the reviewed studies [variable x ] was generally associated with higher / lower rates of [outcome y ].
- A limitation of some / most / all of these studies is [ y ].
Please see this sample annotated literature review from James Mason University.
Structure of a literature review [2]
- Problematization: The 2 to 3 pages of problematization are a distinct, iterative, step. It may take doing such a statement a few times before moving forward to writing the actual paper.
- Search: Write down your keyword sets, your updated keyword sets, and databases. It is perfectly within a reviewer’s rights to ask for these details.
- Summary: Really getting to know major themes requires some annotation of articles. You want to identify core papers and themes and write about them. This helps you really learn the material. [ChatGPT or Wikipedia are no substitute for deep engagement with a paper.]
- Argument: Either outline or create a slide deck that help you express the arguments in your paper. Read them out loud. Have friends look at them. Present them. [Every literature review has an argument. If not, it’s a summary. A summary does not merit publication in a top outlet.]
- Unpacking: Once you’ve nailed the short pitch, unpack the full argument. [ a) Take time in each major section to map out a) the argument, b) the supporting evidence, and the takeaway. b) Take those major sections, reconcile them, make sure they don’t overlap, then move on to writing. c) Sketch out the paper’s sections, tables, figures, and appendices.]
- Writing: Writing is the easy part. You can always put words to the screen. [Revising and improving is hard. Make time to write every day. Improving requires feedback. Find a writing partner to give feedback. Create your tables and figures. Write to them. Make sure the words in the paper align to the visuals.]
- Communicate: When the paper is done, go back and create a paper presentation. [I do this for the papers that I’m most serious about. The act of storyboarding helps me sort out the small pieces of the story that don’t fit together. If I really want it to succeed, I present it. The act of presenting helps me get it right. My best papers sometimes take seven or eight presentations to get it right. Then I return to the paper and fine tune it. Only then, does it have a shot at a top outlet.]
Literature reviews can be published as part of a scholarly article, often after the introduction and sometimes with a header, but they can also be published as a standalone essay. To find examples of what reviews look like in your discipline, choose an appropriate subject database (such as MLA for literary criticism) and conduct a keyword search with the term “Literature Review” added in quotes:
Not only do these examples demonstrate how to structure different types of literature reviews, but some offer insights into trends and directions for future research. In the next section, we’ll take a closer look at some reading strategies to help guide you through this process.
Since scholars already have produced literature reviews on various scholarly conversations, you don’t always need to “reinvent the wheel” (start a literature review from nothing). You can find a published literature review and update it or amend it; scholars do that all the time. However, you must properly cite work you incorporate from others.

| Provide your audience with an overview of the available research on your area(s) of study, including: the literary work, theory, methodology, and method (if the assignment permits). | Skip the literature review. |
| Review only materials about the literary work but not about theory, methodology, and method. | |
| Provide your critical commentary on the materials (if the assignment permits). | Present previous research as though it is all equally good or useful. |
| Build on the research found in other scholarship. | Aim to tear down the research of other scholars. |
- What types of literature review will you be using for your paper? Why did you make this selection over others? If you haven’t made a selection yet, which types are you considering?
- What specific challenges do you face in following a literature review structure?
- If there are any elements of your assignment that need clarification, please list them.
- What was the most important lesson you learned from this page? What point was confusing or difficult to understand?
- In the “Back Matter” of this book, you will find a page titled “Rubrics.” On that page, we provide a rubric for Creating a Literature Review ↵
- Richard West, Brigham Young University, amended by Jason Thatcher, Temple University - https://www.linkedin.com/posts/jason-thatcher-0329764_academicwriting-topten2023-activity-7146507675021766656-BB0O ↵
Writing the Literature Review Copyright © 2021 by Barry Mauer and John Venecek is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License , except where otherwise noted.
Share This Book
- UWF Libraries
Literature Review: Conducting & Writing
- Sample Literature Reviews
- Steps for Conducting a Lit Review
- Finding "The Literature"
- Organizing/Writing
- APA Style This link opens in a new window
- Chicago: Notes Bibliography This link opens in a new window
- MLA Style This link opens in a new window
Sample Lit Reviews from Communication Arts
Have an exemplary literature review.
- Literature Review Sample 1
- Literature Review Sample 2
- Literature Review Sample 3
Have you written a stellar literature review you care to share for teaching purposes?
Are you an instructor who has received an exemplary literature review and have permission from the student to post?
Please contact Britt McGowan at [email protected] for inclusion in this guide. All disciplines welcome and encouraged.
- << Previous: MLA Style
- Next: Get Help! >>
- Last Updated: Aug 8, 2024 11:00 AM
- URL: https://libguides.uwf.edu/litreview
- UNC Libraries
- Course Guides
- ENGL105 - Scholarly Articles 101
Types of Scholarly Articles
Engl105 - scholarly articles 101: types of scholarly articles.
- Peer-review
- Other Types of Sources
Research Articles
Review articles, tips & practice.
- How to Read a Scholarly Article
- Extend Your Knowledge
Sometimes a professor might ask you to find original research or may ask you to not use literature reviews/systematic reviews as sources, but what do those terms mean? How can we tell if our potential source meets our professor's criteria?
In a research article, an original study is conducted by the authors. They collect and analyze data, sharing their methods and results, and then draw conclusions from their analysis. The kind of study performed can vary (surveys, interviews, experiments, etc.), but in all cases, data is analyzed and a new argument is put forth. Research articles are considered primary sources.
- Note: research articles will often contain a section titled "literature review" - this is a section that looks at other existing research as a foundation for their new idea. Simply seeing the words "literature review" does not automatically mean an article is a review article- it is important to look closer
Below is a screenshot of the abstract of the article Effectiveness of Health Coaching in Diabetes Control and Lifestyle Improvement: A Randomized-Controlled Trial , with some words underlined that let us know that a study was conducted and that this is a research article.

A review article gathers multiple research articles on a certain topic, summarizing and analyzing the arguments made in those articles. A review article might highlight patterns or gaps in the research, might show support for existing theories, or suggest new directions for research, but does not conduct original research on a subject. Review articles can be a great place to get an overview of the existing research on a subject. A review article is a secondary source.
- Looking in the reference section of a literature or systematic review can be a good place to find original research studies.
Below is a screenshot of the abstract of the article The Effect of Dietary Glycaemic Index on Glycaemia in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials , with words underlined that clue us in that this is a review article.

Tips for identifying article type
Start by looking at the abstract to determine if a source might be a research article or a review article. If you're not sure after looking at the abstract, find the methods section for the source - what methods did the authors use? If they mention searching databases, it's most likely a review and if they mention conducting an experiment, survey, interview, etc., it's most likely a research article. If you're still unsure, feel free to reach out to a librarian and ask !
Let's Practice
Below are two different scholarly articles. Look at the abstract and the methods section- Which one is an original research study? Which one is a literature review?
- Article 1- Research or Review?
- Article 2- Research or Review?
Research & Instruction Associate

- << Previous: Anatomy of a Scholarly Article
- Next: How to Read a Scholarly Article >>
- Last Updated: Aug 7, 2024 2:00 PM
- URL: https://guides.lib.unc.edu/scholarly-articles-101
Generating knowledge graphs through text mining of catalysis research related literature †

First published on 5th August 2024
Structured research data management in catalysis is crucial, especially for large amounts of data, and should be guided by FAIR principles for easy access and compatibility of data. Ontologies help to organize knowledge in a structured and FAIR way. The increasing numbers of scientific publications call for automated methods to preselect and access the desired knowledge while minimizing the effort to search for relevant publications. While ontology learning can be used to create structured knowledge graphs, named entity recognition allows detection and categorization of important information in text. This work combines ontology learning and named entity recognition for automated extraction of key data from publications and organization of the implicit knowledge in a machine- and user-readable knowledge graph and data. CatalysisIE is a pre-trained model for such information extraction for catalysis research. This model is used and extended in this work based on a new data set, increasing the precision and recall of the model with regard to the data set. Validation of the presented workflow is presented on two datasets regarding catalysis research. Preformulated SPARQL-queries are provided to show the usability and applicability of the resulting knowledge graph for researchers.
Introduction
Information structuring and systematization can be achieved through the utilization of ontologies. An ontology serves as a data model, depicting a collection of concepts and the relationships among these concepts within a specific domain. 7 In this framework, terms are organized hierarchically as classes and subclasses, with each class linked to other classes through properties. This allows for automated classification of research papers with regard to those classes, increasing FAIRness of the classified texts.
Ontology learning (OL) from text is the (semi-)automated process of ontology creation or reuse for enrichment or population purposes. In recent years, several OL approaches have been developed to automate the construction of ontologies. Heuristic and conceptual clustering is one of the statistical-based approaches used for grouping the concepts based on the semantic distance between them to build towards hierarchies. This method was employed in previous work 8 for knowledge extraction from catalysis-related texts for the automatic creation of a taxonomy with important terms extracted from texts. However, the resulting hierarchy is still missing specific interrelations between the terms, and concepts lack proper characterization through axioms. This proves that it is important to integrate the relation extraction in the process of OL. One other tool OntoCmaps 9 is an OL system, with which non-taxonomic relations can be recognized with dependency structure analysis and ontologies are constructed in the form of concept maps, which are not domain-specific and can contain not necessary information.
To extract valuable information from publications in the field of catalysis research, which can be considered as a named entity recognition (NER) task, a pre-trained model, CatalysisIE, 10 was used. This allows for identification of key information from a text based on pre-trained classes, such as names of people. The authors of this model constructed the first benchmark data set for knowledge extraction from the scientific literature in catalysis using active learning to generate a candidate sentence pool for annotation purposes. With this, extracted entities can be categorized into six categories: catalyst, reaction, reactant, product, characterization, and treatment. For the text span representation, pre-trained SciBERT 11 models were used. The parameters of SciBERT were optimized for catalysis-related information extraction (IE) by undergoing the domain adaptation using a corpus consisting of 10.4 million words. 10
The objective of this work is to facilitate the acquisition of information for catalysis research. This is obtained through the design of a tool for the automatic systematization of data extracted from scholarly publications into knowledge graphs. The construction of the knowledge graphs is based on an ontology, which allows for higher data FAIRness by structured relations and conceptual classification of knowledge. Additionally, the content of these publications is preserved in the form of terms deemed relevant to catalysis research. Utilizing CatalysisIE, the extracted entities can be categorized into the six concepts. After preprocessing, the abstracts from scholarly publications can be extracted with natural language processing (NLP) techniques. NLP techniques enable computers to interpret and generate human language, such as scientific texts. For IE, the pretrained model by the authors of CatalysisIE 10 can be used. Furthermore, the CatalysisIE model is trained on the complemented dataset presented in this work.
Ontologies can be queried using SPARQL 12 queries, e.g. formulated in Python functions. SPARQL is a structured query language used to retrieve data stored within databases, especially for triplet-based data, such as ontologies. Automatically generated knowledge graphs containing information for the retrieval of the publications can also later be queried for retrieval of publications.
| Overview of the proposed framework. | ||
Data retrieval
In this work, abstracts were processed assuming that this part of the initial text contains important information about the content of the article and the output is less affected by noisy and repetitive information. The noisy information could be, for example, the previous studies usually mentioned in the introduction section. Abstracts and publication titles were retrieved using text extraction techniques directly from PDF-files. Furthermore, the publicly available CrossRef REST API and publishers' API for metadata retrieval were used. 14 The CrossRef REST API was integrated with the habanero Python package, 15 which fulfils the role of a low-level client that provides functionalities for querying and response handling. The pybliometrics package, 16 an API-wrapper to access Scopus, is used for abstract scraping, which implies the process of automatically mining data or collecting information from the internet, along with pdfdataextractor 17 for abstract extraction directly from PDF files. When an abstract is not able to be retrieved from a PDF, the HTML-version of the publication and thus its abstract are retrieved based on the DOI.
After text preprocessing, entities relevant to the field of catalysis research are extracted from the text using a pre-trained model and labelled with one of the six categories: catalyst, reaction, reactant, product, characterization, and treatment.
Text mining and preprocessing
An example of compound identification with user feedback for “Rh 2 O 3 ” is shown in Fig. 2(a) . In cases where no compounds could be retrieved from PubChem and no synonyms were found in ChEBI, the user is prompted to confirm whether the compound exists. If the user answers “no”, the presumed chemical entity is skipped. By responding “yes”, the user confirms the existence of the entity and it will be identified with the name provided to the user (see Fig. 2(b) ).
| Example of a user-feedback process during preprocessing in Python. Identification of chemical compounds using queried compounds from PubChem (a) and confirmation of the compound existence (b). User input is marked in green. | ||
Ontology learning
As the main working ontology, the Allotrope Foundation Ontology (AFO) 22 is selected. In this ontology, object properties important for the ontology extension process are created manually ( Table 4 in the appendix), while classes and data properties are created automatically. After preprocessing of the extracted entities based on their labelled roles, the working ontology is extended with classes and instances using existing terms in other ontologies. Furthermore, rule-based approaches in combination with syntactic dependency parsing are pursued. The initial preprocessing of the extracted entities includes the POS taggers, lemmatization, tokenization, and the use of regular expressions. In specific steps, such as lemmatization, modules from spaCy 23 are used. Thus, the preprocessing allows prevention of the creation of synonyms as independent concepts in the ontology and creation of relations based on the context in the text. For example, given an entity “RhCo/SiO 2 ” labelled “catalyst”, “RhCo” is recognised as a catalyst, which is supported on “SiO 2 ”. This relation will be added later in the ontology to the corresponding classes of the compounds. To recreate the hierarchical structure within the ontology, syntactic dependency parsing was utilized as a preprocessing step to define the “head” – word of the entity and use it for search in ontologies. For example, the “head” of “packed bed methanation” is “methanation”.
In this work, some existing ontologies were used in the workflow for ontology extension. The primary selection criterion for these ontologies was their relevance to the domain of catalysis research. The selection of ontologies for this work, which are listed along with a short description in Table 1 , is based on the overview from the collection of ontologies for catalysis research presented in ref. 24 .
| Ontology acronym | Full name | Description |
|---|---|---|
| BFO | Basic Formal Ontology | A small ontology is designed for use in analysis and integration in scientific or other domains. It does not contain physical, chemical, or other domain-specific terms and is well used as a top-level ontology |
| AFO | Allotrope Foundation Ontology | AFO is a domain-specific ontology, offering a standardized vocabulary and semantic framework for the representation of laboratory analytical processes. AFO is aligned with BFO as a top-level ontology. Reasoning can be only provided by the HermiT reasoner |
| ChEBI | Chemical Entities of Biological Interest | ChEBI represents a vocabulary with a focus on small molecular entities and contains such information as InChiKeys, CAS numbers, and exact and related synonyms of chemical compounds |
| MOP | Molecular Process Ontology | Domain ontology contains good conceptual descriptions of molecular processes, such as crystallization and methylation |
| RXNO | Name Reaction Ontology | Domain ontology is strongly connected to MOP. It contains more than 500 classes representing organic reactions with good conceptual description |
Ontology extension is based on the reuse of the classes existing in other ontologies to properly characterize concepts and reuse existing frameworks and axioms. The reuse of ontology terms creates links between data, making the ontology more valuable. 28
For example, ChEBI contains more than 3 million axioms. Thus, only relevant subsets of ontologies were reused. After searching for classes within the working ontology using the owlready2 Python package, 29 missing chemical compounds and possible reaction types are searched for in other ontologies as listed in Table 1 . To accomplish this, a nested dictionary is used, containing all IRIs of terms alongside their corresponding labels, prefLabels, altLabels, and the names used within the ontology. Applying a class information extraction process that incorporates functions from the owlready2 package, the dictionary was generated for 22 ontologies relevant to the domain of catalysis research. 24 Once the dictionary is loaded into the Python environment, it is searched for classes missing in the working ontology. If one of the labels, prefLabels, altLabels, or names matches the searched entity, the corresponding IRI is added to the dictionary along with the matching value. The IRIs of found terms that are still missing in the working ontology are stored in automatically created text files. The names of the text files include the acronym of the source ontology for further reference.
Ontology extension
This ensures that the module retains all the same logical entailments in the full ontology, providing consistency in the ontology subset. The chosen SMLE approach is the BOTTOM module, which contains the terms in the seed, their corresponding superclasses, and the interrelations between them. As the name implies, the class hierarchy is built from the bottom up, gathering the superclasses of the selected class. Thus, for each ontology, a separate subset of relevant classes is created in rdf/xml (owl) format.
The second task for which ROBOT is used in this work is to merge the created subsets of classes and the main ontology into a single ontology with a single .owl-file. Thus, the merging process is used to update the working ontology within existing terms of other ontologies.
Because some of the merged ontologies are aligned with different top-level ontologies, terms that theoretically share the same definition are located at different positions within the class hierarchy. For example, the OBO is the top-level ontology of ChEBI, while the AFO is aligned with the BFO. Both ontologies have, for example, the term “atom”, but at different positions in the class hierarchy. Another factor why the same terms are represented differently is related to the granularity problem of ontologies. This issue arises because ontologies often adopt different levels of details when representing identical knowledge to support different applications. 30
Since all of the utilized ontologies are connected to the domain of catalysis 24 and chemistry, 31 terms with identical designation are assumed equivalent. The equivalence of classes indicates that respective classes share all their instances, and the descriptions of both classes are interlinked. However, the use of the equivalence relation does not imply class equality. Both relations are defined differently in OWL. Equality is denoted by “owl:sameAs”, while the equivalence is represented by owl:equivalentClass. Class equality can only be defined by the description language OWL-Full, and owlready2 supports only equivalence. 29
To identify terms with the same designation that originate from different ontologies and consequently have different IRIs, the mappings created in previous work 24 are used in the processing. These mappings represent all terms shared by two ontologies according to the same IRIs or the same set of labels, prefLabels, names, or altLabels. After merging ontologies to reuse existing terms, the process of creating new classes and subsequently populating the working ontology with new instances is initialized. First, a new instance of a publication is generated as an instance of the “publication” superclass. The DOI and title of the publication retrieved at the beginning of the process are added to the publication instance as datatypes using the “has doi” and “has title” datatype properties, respectively. Extracted chemical compounds that do not exist in the working ontology after merging are then created as new classes within the working ontology, utilizing the context information of the new compounds. Chemical compounds that can be further broken down into compounds and atoms, such as “Al 2 O 3 ” or “titanium dioxide”, or those that are recognized as compounds using pubchempy are created as subclasses of the “molecule” class.
Support material entities, which represent a combination of two or more carrier compounds, like “TiO 2 –SiO 2 ”, or materials such as “MCM-41” are created as subclasses of the “support material” class. Each newly created class and instance are automatically assigned a generated name linked to the number of the processed publications in the working ontology.
Entities from the “Reactant”, “Product” and “Catalyst” categories that represent specific types of chemical entities, such as “light olefin” and “vapour phase propene”, are created as instances of the corresponding chemical compound. Extracted and preprocessed catalyst entities are created as instances of the “chemical substance” class.
Chemical entities, which represent catalysts in the form of
“ <catalytic compound>/<support compound> ” or
“ <catalytic compound>@<support compound> ”
are labelled in the ontology
“ <catalytic compound> supported on <support compound> ”
and linked with their chemical compounds based on their roles in the entity using the “catalytic component of” and “support component of” object properties. A schematic example of interconnections within the ontology is shown in Fig. 3 .
| Examples of created entities and assigned relations. Entities within dashed boxes represent instances, while continuous bordered boxes represent classes. | ||
Table 4 in the appendix lists the object properties and their inverse properties that need to be defined within the working ontology in order to assign the relations between the newly created entities.
The creation of the classes corresponding to the catalyst types is based on the creation of subclasses of the term “catalyst role”, which already exists in the AFO ontology. Roles in ontologies are used to reduce the amount of object properties and thus to speed up reasoning. The corresponding roles of terms are provided as classes in the ontology and terms are linked to them via the “has role” object property. The hierarchical structuring of the catalyst roles is based on the content of the classes extracted from the entities. For instance, within the text corpus, the extracted class of an entity after preprocessing might be a “dispersed catalyst role”, while the catalyst type corresponding to another entity is an “atomically dispersed catalyst role”. Since the second class is identical to the first but with an additional word, it is considered a subclass of the first class. In case no entity from the text corpus has a “dispersed catalyst role” as an extracted class, then an “atomically dispersed catalyst role” is created as a subclass of the “catalyst role”.
Chemical reactions are created as subclasses of the previously extracted reaction “heads”. If there is no corresponding reaction found in other ontologies, a new class is created as a subclass of “chemical reaction (molecular)”, which is also a class within the AFO. For each class created after the merging that corresponds to an extracted entity or a chemical compound, an instance is created with an automatically generated name.
The label of the instance is the same as the label of its corresponding class. The same procedure is applied to the newly created classes. All classes and instances, once created, can be reused for ontology extension with the respective publications. The newly created classes of chemical compounds are linked to their corresponding components via “has part” relations at the instance level.
Created instances are linked to their roles according to the categories and the context using the “has role” relation. The used roles include the “support role”, “reactant role”, “product role”, and “catalyst role”, and all are created as subclasses of the “catalyst role”.
Finally, all created and used instances that are mentioned in the processed publication are linked to the instance of the publication through the “mentioned in” object property. Entities labelled “Characterization” and “Treatment” are added to annotations of publications as comment.
SPARQL queries
The following competency questions were implemented as SPARQL queries and can thus be easily retrieved from the knowledge graph resulting from the extension of the ontology. The corresponding SPARQL queries are numbered and exemplary input and output of the queries are listed in Table 5 in the appendix and an exemplary SPARQL query is listed in Table 6 in the appendix:
• Give me a list of reactions (1), reactants, support materials, catalysts, and products mentioned in one specific publication, which is a part of the knowledge graph, in one list (2) or separately,
• Retrieve the abstracts from publications in the ontology (3),
• Give me a list of DOIs of publications from the working ontology, which mention the same reactions (4) or the specific reaction (5) or catalyst (6),
• Give me a list of reactions, reactants, support materials, catalysts, and products mentioned in all publications of the knowledge graph (7),
• I need a list of all possible synonyms for the extracted reactants (8), support materials (9), catalysts (10), and products (11) in the form of chemical entities,
• I need possible catalysts where the support material from this paper can be used (12).
The retrieved entities can be used to query Scopus for new publications with similar context. Using the pybliometrics Python package, the search is performed, leading to a query, which has the same structure as a query that works in the Scopus advanced search. With the chosen query type ‘TITLE-ABS-KEY()’ (as depicted exemplarily in Fig. 4 ), the search is performed within the titles, abstracts, and keywords of the publication.
| Two types of query formulations for the advanced search for further publications in Scopus, executed by the Python API. | ||
Since there are multiple ways to name a specific chemical compound, to avoid a large number of possible queries and at the same time allow diversity in the naming of chemical compounds, a trade name or common name and a formula listed in the class annotations of a chemical compound are used for queries' formulation. Moreover to exclude mismatches, the publication will be skipped if during text mining no reaction was found in the text. After a query is executed, its results are downloaded and cached to speed up the subsequent analysis.
After the results are concatenated into one table, duplicates are removed from it. As Scopus contains records of articles published since 1970, an option to filter the results by publication date is integrated into the process, to allow for the inclusion of primarily newer publications into the knowledge graph. Utilization of the pandas Python module 33 allows the resulting DataFrames to be stored as sheets in an Excel file.
Results and discussion
Moreover, a set of 28 publications on methanation processes (dataset 2) was used to evaluate how well the created tool works on the different types of catalysed reactions. Hereby the focus was laid on the heterogeneously catalysed conversion of carbon monoxide and carbon dioxide to methane via hydrogenation, which is important for the production of synthetic natural gas. In particular, the valorization of CO 2 together with renewable hydrogen might be considered an integral sustainable path towards the production of renewable gaseous fuels. 36 For that, an extension of an alternate ontology setup similar to the first dataset was performed.
The dataset for training of the model was complemented with 151 sentences manually labelled in label-studio 37 from 18 abstracts of papers to the topic of hydroformylation in the liquid and gas phase. Checkpoints from the model trained by the authors of CatalysisIE and the model trained on the complemented dataset were compared with each other.
To evaluate the difference in the prediction of the checkpoints, ten manually labelled abstracts from papers to the same topic were compared to predictions of both models. Since it is important to gain as many correct distinct predictions from the text as possible to be able to describe the content of the publication using extracted entities, the recall R of the model was evaluated with the number of true positives TP and false negatives FN using eqn (1) . To obtain the true positives and false negatives, the amount of distinct entities was counted and compared with the number of distinct entities from the prediction after qualitative manual labelling of the texts. This comparison for each extracted abstract from dataset 1 is shown in Table 9 in the appendix.
Besides recall, the precision Pr was selected for evaluation of multi-label classification. Because class imbalances are present in the dataset, the precision was calculated using eqn (2) with the number of positives P i instead of true positives TP and the number of used labels N . Furthermore, the standard deviation σ of the precision was selected as a metric and calculated using eqn (3) . The sum of true positives corresponds to the number of correctly predicted instances. Precision and its standard deviation were calculated for the six categories for each of the abstracts.
| (1) |
| (2) |
| (3) |
Extraction of sequences was treated as a binary classification problem, where the sum of TP is correctly extracted from distinct entities and is independent from the assigned label. The sum of true positives and false negatives is the total number of distinct manually labelled entities in the text. The metrics were calculated for the ten manually labelled abstracts from papers to the same topic and compared to predictions of both checkpoints I and II. Here, checkpoint I addresses the complemented model, while checkpoint II addresses the pre-trained checkpoint, provided by the developers of CatalysisIE. The resulting metrics are listed in Table 2 . Deviations in the metrics of the fourth publication may be due to formatting errors in the retrieved abstract, causing extracted tokens to end with citation numbers ( e.g. , “catalysts9”), thus not being counted as found entities.
| TP + FN | CP | TP | R | Pr | σ | |
|---|---|---|---|---|---|---|
| 1 | 12 | I | 11 | 91.7 | 83.3 | 40.8 |
| II | 10 | 83.3 | 66.7 | 51.6 | ||
| 2 | 15 | I | 14 | 93.3 | 93.3 | 14.9 |
| II | 12 | 80.0 | 80.0 | 44.7 | ||
| 3 | 15 | I | 13 | 86.7 | 75.6 | 43.3 |
| II | 12 | 70.0 | 73.3 | 43.5 | ||
| 4 | 16 | I | 6 | 37.5 | 35.0 | 23.8 |
| II | 6 | 37.5 | 38.7 | 30.7 | ||
| 5 | 15 | I | 10 | 66.7 | 64.6 | 9.9 |
| II | 7 | 66.0 | 61.8 | 26.7 | ||
| 6 | 15 | I | 13 | 86.7 | 58.3 | 49.2 |
| II | 14 | 93.3 | 66.7 | 51.6 | ||
| 7 | 37 | I | 33 | 89.2 | 88.2 | 21.7 |
| II | 24 | 64.9 | 62.7 | 16.3 | ||
| 8 | 29 | I | 25 | 86.2 | 68.2 | 10.5 |
| II | 24 | 82.8 | 64.7 | 11.4 | ||
| 9 | 25 | I | 12 | 48.0 | 63.6 | 26.0 |
| II | 13 | 52.0 | 68.6 | 25.1 | ||
| 10 | 6 | I | 6 | 100.0 | 100.0 | 0.0 |
| II | 6 | 100.0 | 100.0 | 0.0 |
The entities labelled “Characterization” were predicted least accurately. Additionally, there were no “Treatment” labels in the evaluation dataset. Overall, the model trained on the expanded dataset (CP I) was better at predicting entities labelled “Catalyst”. The average recall of the newly trained model for ten abstracts is equal to 86.67% with a standard deviation of 20.85% and shows a high average precision of 71.90%. In comparison, the recall of the old model (CP II) is 80.00% with a standard deviation of 19.37% and an average precision of 66.67%. In both cases only in one text, precision and recall fall under 50%. Furthermore, for the ten publications shown in Table 2 , in the cases where CP I achieved higher precision, σ was lower. This indicates that the dispersion across the different classes in relation to Pr has decreased and therefore the model makes more stable predictions across the classes.
To investigate the performance of the extended model further, ten abstracts from dataset 2 are labelled manually and classified with CP I and CP II to evaluate the metrics as in Table 2 . The resulting metrics are presented in more detail in Table 7 . An average recall of 82.81% with a standard deviation of 22.39% and an average precision of 71.46% was achieved for CP I. Furthermore, an average recall of 79.47% with a standard deviation of 19.92% and an average precision of 73.20% was achieved for CP II. Thus, the extended model can also be applied on dataset 2.
Title recognition by 19 out of 23 processed PDFs from dataset 1 was successful and 26 from 28 publications from dataset 2 could be recognized correctly. Publications of “Royal Society of Chemistry” could not be correctly recognized because the layout of the publications is not integrated in the workflow of the used pdfdataextractor package.
The AFO was chosen as the initial ontology, because of its linkage to the chemical domain and well-defined structure in the class hierarchy. Table 8 lists the terms and textual definitions assigned as equivalent in ontologies for both datasets, which exist in the AFO and are merged into the working ontology from ChEBI.
Chemicals which could not be found in PubChem or in ChEBI are created as instances of the class “chemical substance”. For dataset 1, the ontology is extended with 53 instances of “chemical substance”. Dataset 2 results in 55 instances of “chemical substance” that were also created automatically. Each of the generated instances representing extracted entities and their chemical components is provided with a connection to the publication in which it is mentioned and linked to the corresponding roles as shown in an excerpt of the resulting ontology in Fig. 5 . The reactions that are mentioned within the publication are listed, including the respective participants of the reactions within the knowledge base (upper area of the figure). The individual “cobalt atom”, for example, is connected with the individual “Co-containing catalyst” via the object property “catalytic component of” (right area of the figure), thus indicating the suitable catalytic component of the concept extracted from text. Furthermore, the role of a “bimetallic catalyst role” is asserted to the three individuals on the bottom right of Fig. 5 . The class “bimetallic catalyst role” is created as a subclass of the “catalyst role”, which in turn also has an individual that is connected to other substances via the “has role” object property (bottom left of the figure).
| Excerpt from the created ontology for dataset 1 created with Protégé. Boxes marked with yellow circles represent classes and those with purple rhombi are instances. Arrows denote the relationships between them, color-coded as listed in the legend on the right. Small boxes with a plus (+) inside indicate that not all relations of the entity are shown in the figure. | ||
The knowledge graph with publications from dataset 1 was extended by 48 classes from the other ontologies, including their superclasses and interrelations. In total, 331 new classes, 9 new object properties, 2 new data properties, and 155 new individuals were added to the working ontology. From the new classes, 288 were merged from other ontologies, while none of the new individuals were merged from other ontologies, as expected. The new object and data properties were merged from other ontologies.
In the knowledge graph with dataset 2, 39 classes from other ontologies were imported from other ontologies together with their respective superclasses and interrelations. With this, 222 new classes, 4 new object properties, 2 new data properties, and 130 new individuals were added to the working ontology. Here, 198 from the 222 new classes were merged from other ontologies, while also none of the new individuals were merged from other ontologies. The new object and data properties were merged from the other ontologies listed in Table 1 and counted without the ones already presented in Table 4 in the appendix. The explained ontology metrics are listed in Table 3 .
| Metric | Initial ontology | Extended ontology dataset 1 | Extended ontology dataset 2 |
|---|---|---|---|
| Classes | 3116 | 3447 | 3338 |
| Instances | 47 | 203 | 178 |
| Logical axioms | 5755 | 6936 | 6596 |
| SubClassOf | 4823 | 5372 | 5174 |
| Equivalent classes | 178 | 188 | 185 |
Fig. 6 shows the individual “0.5% Co–0.5% Rh supported on Al 2 O 3 ” in an excerpt of Protégé after reasoning with HermiT. 38 The implicit knowledge is highlighted in yellow, showing an increased semantic expressiveness for the individual describing the catalyst complex. Thus, the individual now also can be found when searching the knowledge graph, e.g. , for catalysts that contain cobalt.
| Excerpt from Protégé with inferred relations after reasoning for the individual “0.5% Co–0.5% Rh supported on Al O ”. Knowledge inferred by the reasoner is highlighted in yellow, showing increased semantic expressiveness of the individual. | ||
Most of the terms in both knowledge graphs originate from the ChEBI ontology and identify chemical compounds and atoms. But also, the classes for such terms as “hydrogenation”, “hydroformylation”, and “acylation” are reused from the RXNO ontology. In the current process, entities representing some chemical groups, such as “phenolic substances” or “phenolic species”, can be recognized with the text mining module, but the extension of the ontology with them is not implemented. This includes, for example, entities such as “phenolic substances”, “phenolic species”, and “alkyl species” which are usually classified as products or reactants in the text. Such entities cannot be queried in PubChem, and in ChEBI, the presumed superclasses are placed in different positions. All queries are formulated within the functions in the module “queries” provided in the GitHub repository of this work 39 and can be executed by the Jupiter notebook “user_queries.ipynb” contained in the repository. It contains descriptions of code cells, which execute specific queries, which can answer competency questions formulated in Methods. In addition, some examples of executed functions are also provided in the notebook.
Listings of the resulting publications are found in the provided GitHub repository of this work 39 in the “output” directory.
To further rate the quality of the query result, a random sample in size of 50 publications from the resulting filtered list of 731 publications similar to those of dataset 1 was selected for the evaluation of the queried content. The list of chosen publications for evaluation is provided in the appendix in Table 9 .
These publications are rated as similar to the publications in the knowledge graph (of dataset 1) if the following requirements are fulfilled, based on the evaluation of their abstracts and titles:
• Heterogeneous or homogeneous catalysis or catalysts are mentioned.
• Hydroformylation or hydrogenation is mentioned.
• Rh-, Co-, and Ni-based catalysts with silica, zeolite or aluminium oxide as a support material are mentioned.
According to these restrictions, 34 out of 50 publications of the sample provided are rated as similar to the content of the publications within the knowledge graphs, which is equal to 68% accuracy.
Conclusions
The quality of matched articles to the content of the knowledge graphs was evaluated with dataset 1, dealing with hydroformylation reactions. A random sample of 50 publications was investigated in more detail and abstracts, keywords, and titles were screened for the mention of catalysts, hydrogenation or hydroformylation, and whether Rh-, Co-, and Ni-based catalysts with silica, zeolite or aluminium oxide as a support material were mentioned. With these quite strict criteria and with the lack of case sensitivity in the Scopus API, 68% accuracy was achieved for the random sample. This allows for more structured searches of relevant scientific literature in the domain of catalysis research, which is highly important, especially in this domain, as research is quite heterogeneous and the number of relevant publications in the field is quite high. However, a more thorough post-processing of these found publications needs to be conducted, e.g. by a post-processing that conducts a case-sensitive automated search of the respective keywords in the extracted abstracts from Scopus, to improve the accuracy of the output related publications.
| Object property | Reverse object property | ||
|---|---|---|---|
| Name | Rdfs:Label | Name | Rdfs:Label |
| supported_on | Supported on | support_material_of | Support material of |
| support_component_of | Support component of | has_support_component | Has support component |
| catalytic_component_of | Catalytic component of | has_catalytic_component | Has catalytic component |
| mentioned_in | Mentioned in | Mentions | Mentions |
| Query no. | Input parameters | Output |
|---|---|---|
| 1 | Doi = r‘10.1021/acsami.0c21749.s001’ | [‘hydroformylation’] |
| 2 | list_type = ‘all’ | [‘hydroformylation’, ‘olefin’, ‘rhodium atom’, ‘Rh-based atomically dispersed catalyst’, ‘Rh supported on ZnO modified with Pi’, ‘zinc oxide’, ‘phosphate ion’, ‘aldehyde’, ‘linear aldehyde’] |
| Doi = r‘10.1021/acsami.0c21749.s001’ | ||
| 3 | Doi = r‘10.1021/acsami.0c21749.s001’ | Abstract: in the study of heterogeneity of homogeneous processes, effective control of the microenvironment of active sites… |
| 4 | Doi = r‘10.1021/acsami.0c21749.s001’ | [[‘10.1021/acscatal.1c02014.s001’], [‘10.1021/acscatal.0c04684.s001’], [‘10.1021/acscatal.1c00705.s002’], [‘10.1021/acscatal.1c04359’],…] |
| 5 | Reac = “hydrogenation”, Doi = none | [[‘10.1021/acscatal.0c04684.s001’], [‘10.1021/acscatal.1c00705.s002’],…] |
| 6 | Cat = “RhCo”, Doi = none | [[‘10.1021/acscatal.0c04684.s001’], [‘10.1021/acscatal.1c00705.s002’], |
| 7 | list_type = ‘all’ | [‘styrene’, ‘cobalt atom’, ‘0.5% Co–0.5% Rh supported on Al O ’, ‘Co-containing catalyst’, ‘aluminum oxide’, ‘hydroformylation’,…] |
| Doi = none | ||
| 8 | list_type = ‘reactant’ | [‘olefin’] |
| Doi = r‘10.1021/acsami.0c21749.s001’ | ||
| 9 | list_type = ‘product’ | [‘Aldehyd’, ‘RCHO’, ‘aldehidos’, ‘aldehydes’, ‘Aldehyde’, ‘aldehydum’, ‘an aldehyde’, ‘RC( |
| Doi = r‘10.1021/acsami.0c21749.s001’ | ||
| 10 | Doi = r‘10.1021/acsami.0c21749.s001’ | [[‘45Rh’, ‘rhodium’, ‘Rh’, ‘rodio’, ‘Rh(111)’, ‘rhodium atom’]], [[‘Rh on ZnO modified with Pi’, ‘Rh supported on ZnO modified with Pi’]] |
| 11 | Doi = r‘10.1021/acsami.0c21749.s001’ | [[‘zinc oxide’, ‘oxyde de zinc’, ‘Zinkoxid’, ‘oxido de cinc’, ‘ZnO’], [‘phosphate ions’, ‘Pi’, ‘phosphate’, ‘phosphate ion’]] |
| 12 | Doi = r‘10.1021/acscatal.1c02014.s001’ | [[‘silicon dioxide’, ‘Rh supported on SiO catalyst’], [‘silicon dioxide’, ‘Rh P nanoparticle supported on SiO support material’], [‘silicon dioxide’, ‘Rh7Co1P4 supported on SiO ’],…] |
| only_doi = false |
| TP + FN | CP | TP | R | Pr | σ | |
|---|---|---|---|---|---|---|
| 1 | 15 | I | 13 | 86.7 | 68.3 | 32.5 |
| II | 12 | 80.0 | 61.7 | 36.1 | ||
| 2 | 19 | I | 15 | 78.9 | 82.7 | 28.9 |
| II | 15 | 78.9 | 82.7 | 28.9 | ||
| 3 | 9 | I | 9 | 100.0 | 93.3 | 14.9 |
| II | 9 | 100.0 | 93.3 | 14.9 | ||
| 4 | 11 | I | 6 | 54.5 | 66.7 | 28.9 |
| II | 7 | 63.6 | 70.8 | 26.0 | ||
| 5 | 6 | I | 6 | 100.0 | 100.0 | 0.0 |
| II | 6 | 100.0 | 100.0 | 0.0 | ||
| 6 | 14 | I | 8 | 57.1 | 43.8 | 51.5 |
| II | 8 | 57.1 | 43.8 | 51.5 | ||
| 7 | 12 | I | 12 | 100.0 | 73.6 | 29.7 |
| II | 11 | 91.7 | 72.6 | 42.6 | ||
| 8 | 12 | I | 7 | 58.3 | 69.3 | 41.3 |
| II | 6 | 50.0 | 49.3 | 46.6 | ||
| 9 | 9 | I | 4 | 44.4 | 69.0 | 27.0 |
| II | 5 | 55.6 | 73.8 | 25.1 | ||
| 10 | 15 | I | 15 | 100.0 | 92.6 | 13.5 |
| II | 15 | 100.0 | 92.6 | 13.5 |
| Term label | IRIs + Definitions | |
|---|---|---|
| In the AFO | In ChEBI | |
| ‘Chemical substance’ | http://purl.allotrope.org/ontologies/material#AFM_0001097 | http://purl.obolibrary.org/obo/CHEBI_33250 |
| A chemical substance is a portion of material that is matter of constant composition best characterized by the entities (molecules, formula units, atoms) it is composed of [IUPAC] | A chemical entity constituting the smallest component of an element having the chemical properties of the element | |
| ‘Anion’ | http://purl.allotrope.org/ontologies/material#AFM_0000161 | http://purl.obolibrary.org/obo/CHEBI_22563 |
| An anion (−) is an ion with more electrons than protons, giving it a net negative charge (since electrons are negatively charged and protons are positively charged) | A monoatomic or polyatomic species having one or more elementary charges of the electron | |
| ‘Ion’ | http://purl.allotrope.org/ontologies/material#AFM_0000077 | http://purl.obolibrary.org/obo/CHEBI_24870 |
| An ion is an atom or molecule in which the total number of electrons is not equal to the total number of protons, giving the atom or molecule a net positive or negative electrical charge | A molecular entity having a net electric charge | |
| ‘Role’ | http://purl.obolibrary.org/obo/BFO_0000023 | http://purl.obolibrary.org/obo/CHEBI_50906 |
| B is a role means: b is a realizable entity and b exists because there is some single bearer that is in some special physical, social, or institutional set of circumstances in which this bearer does not have to be and b is not such that, if it ceases to exist, then the physical make-up of the bearer is thereby changed [BFO] | A role is particular behavior which a material entity may exhibit | |
| ‘Cation’ | http://purl.allotrope.org/ontologies/material#AFM_0000189 | http://purl.obolibrary.org/obo/CHEBI_36916 |
| A cation (+) is an ion with fewer electrons than protons, giving it a positive charge | A monoatomic or polyatomic species having one or more elementary charges of the proton | |
| ‘Group’ | http://purl.obolibrary.org/obo/BFO_0000023 | http://purl.obolibrary.org/obo/CHEBI_24433 |
| A group is an aggregate of people | A defined linked collection of atoms or a single atom within a molecular entity | |
| ‘Atom’ | http://purl.allotrope.org/ontologies/material#AFM_0001028 | http://purl.obolibrary.org/obo/CHEBI_33250 |
| An atom is a smallest particle still characterizing a chemical element. It consists of a nucleus of a positive charge carrying almost all its mass (more than 99.9%) and Z electrons determining its size | A chemical entity constituting the smallest component of an element having the chemical properties of the element | |
| ‘Chemical substance’ | http://purl.allotrope.org/ontologies/material#AFM_0001097 | http://purl.obolibrary.org/obo/CHEBI_59999 |
| A chemical substance is a portion of material that is matter of constant composition best characterized by the entities (molecules, formula units, atoms) it is composed of | A chemical substance is a portion of matter of constant composition, composed of molecular entities of the same type or of different types | |
Data availability
Author contributions, conflicts of interest, acknowledgements.
- D. W. Hook, S. J. Porter and C. Herzog, Dimensions: Building Context for Search and Evaluation, Front. Res. Metr. Anal. , 2018, 3 DOI: 10.3389/frma.2018.00023 .
- M. D. Wilkinson, et al. , The FAIR Guiding Principles for scientific data management and stewardship, Sci. Data , 2016, 3 , 160018, DOI: 10.1038/sdata.2016.18 .
- A. Salazar, B. Wentzel, S. Schimmler, R. Gläser, S. Hanf and S. A. Schunk, How Research Data Management Plans Can Help in Harmonizing Open Science and Approaches in the Digital Economy, Chemistry , 2023, 29 (9), e202202720, DOI: 10.1002/chem.202202720 .
- B. V. Elsevier, Scopus, 2024. Accessed: February 2024. [Online]. Available: https://www.scopus.com/.
- C. P. Marshall, J. Schumann and A. Trunschke, Achieving Digital Catalysis: Strategies for Data Acquisition, Storage and Use, Angew. Chem., Int. Ed. , 2023, 62 (30), e202302971, DOI: 10.1002/anie.202302971 .
- M. Suvarna, A. C. Vaucher, S. Mitchell, T. Laino and J. Pérez-Ramírez, Language models and protocol standardization guidelines for accelerating synthesis planning in heterogeneous catalysis, Nat. Commun. , 2023, 14 (1), 7964, DOI: 10.1038/s41467-023-43836-5 .
- S. Mishra and S. Jain, A Study of Various Approaches and Tools on Ontology, in 2015 IEEE International Conference on Computational Intelligence & Communication Technology , Ghaziabad, India, 2015, pp. 57–61 Search PubMed .
- A. S. Behr, M. Völkenrath and N. Kockmann, Ontology extension with NLP-based concept extraction for domain experts in catalytic sciences, Knowl. Inf. Syst. , 2023, 65 (12), 5503–5522, DOI: 10.1007/s10115-023-01919-1 .
- A. Zouaq, D. Gasevic and M. Hatala, Towards open ontology learning and filtering, Information Systems , 2011, 36 (7), 1064–1081, DOI: 10.1016/j.is.2011.03.005 .
- Y. Zhang, C. Wang, M. Soukaseum, D. G. Vlachos and H. Fang, Unleashing the Power of Knowledge Extraction from Scientific Literature in Catalysis, J. Chem. Inf. Model. , 2022, 62 (14), 3316–3330, DOI: 10.1021/acs.jcim.2c00359 .
- I. Beltagy, K. Lo and A. Cohan, SciBERT: A Pretrained Language Model for Scientific Text, EMNLP. [Online]. Available: https://arxiv.org/pdf/1903.10676.pdf.
- W3C Sparql 1.1. [Online]. Available: https://www.w3.org/TR/sparql11-update/.
- J. Hastings, et al. , ChEBI in 2016: Improved services and an expanding collection of metabolites, Nucleic Acids Res. , 2016, 44 (D1), D1214–D1219, DOI: 10.1093/nar/gkv1031 .
- CrossRef, CrossRef API Documentation. Accessed: 2024.
- S. Chamberlain, J. Maupetit, S. Peak, C. Talbert, D. Himmelstein and K. Niemeyer, Habanero: Python client for the Crossref API, 2024, Accessed: 2024. [Online]. Available: https://github.com/sckott/habanero.
- M. E. Rose and J. R. Kitchin, pybliometrics: Scriptable bibliometrics using a Python interface to Scopus, SoftwareX , 2019, 10 , 100263, DOI: 10.1016/j.softx.2019.100263 .
- M. Zhu and J. M. Cole, PDFDataExtractor: A Tool for Reading Scientific Text and Interpreting Metadata from the Typeset Literature in the Portable Document Format, J. Chem. Inf. Model. , 2022, 62 (7), 1633–1643, DOI: 10.1021/acs.jcim.1c01198 .
- M. C. Swain and J. M. Cole, ChemDataExtractor: A Toolkit for Automated Extraction of Chemical Information from the Scientific Literature, J. Chem. Inf. Model. , 2016, 56 (10), 1894–1904, DOI: 10.1021/acs.jcim.6b00207 .
- Python Software Foundation, re - Regular expression operations, 2024.
- S. Kim, et al. , PubChem 2023 update, Nucleic Acids Res. , 2023, 51 (D1), D1373–D1380, DOI: 10.1093/nar/gkac956 .
- M. Swain, PubChemPy: A way to interact with PubChem in Python, 2014, [Online]. Available: https://github.com/mcs07/PubChemPy.
- Allotrope Foundation, Allotrope Foundation Ontologies. Accessed: 2022.
- I. Montani et al. , spaCy: Industrial-strength Natural Language Processing in Python, 2022.
- A. S. Behr, H. Borgelt and N. Kockmann, Ontologies4Cat: investigating the landscape of ontologies for catalysis research data management, J. Cheminf. , 2024, 16 (1), 16, DOI: 10.1186/s13321-024-00807-2 .
- R. Arp, B. Smith and A. D. Spear, Building ontologies with Basic Formal Ontology , Massachusetts Institute of Technology, Cambridge, Massachusetts, 2015 Search PubMed .
- C. Batchelor, Molecular Process Ontology (MOP). [Online]. Available: https://github.com/rsc-ontologies/rxno.
- C. Batchelor, Chemical Reactions Ontology (RXNO). [Online]. Available: https://github.com/rsc-ontologies/rxno.
- R. C. Jackson, J. P. Balhoff, E. Douglass, N. L. Harris, C. J. Mungall and J. A. Overton, ROBOT: A Tool for Automating Ontology Workflows, BMC Bioinf. , 2019, 20 (1), 407, DOI: 10.1186/s12859-019-3002-3 .
- J.-B. Lamy, Owlready: Ontology-oriented programming in Python with automatic classification and high level constructs for biomedical ontologies, Artif. Intell. Med. , 2017, 80 , 11–28, DOI: 10.1016/j.artmed.2017.07.002 .
- P. Sun and S. Zhang, Identifying Granularity Differences between Large Biomedical Ontologies through Rules, AMIA Annu. Symp. Proc. , 2010, 2010 , 927–931 Search PubMed .
- P. Strömert, J. Hunold, A. Castro, S. Neumann and O. Koepler, Ontologies4Chem: the landscape of ontologies in chemistry, Pure Appl. Chem. , 2022, 94 (6), 605–622, DOI: 10.1515/pac-2021-2007 .
- SPARQL 1.1 Query Language , ed. E. Prud'hommeaux, S. Harris and A. Seaborne, W3C, 2013, [Online] Available: https://www.w3.org/TR/sparql11-query Search PubMed .
- W. McKinney, Data Structures for Statistical Computing in Python, in Proceedings of the 9th Python in Science Conference , Austin, Texas, 2010, pp. 56–61 Search PubMed .
- Y. Liu, et al. , Rhodium nanoparticles supported on silanol-rich zeolites beyond the homogeneous Wilkinson's catalyst for hydroformylation of olefins, Nat. Commun. , 2023, 14 (1), 2531, DOI: 10.1038/s41467-023-38181-6 .
- S. Hanf, L. Alvarado Rupflin, R. Gläser and S. Schunk, Current State of the Art of the Solid Rh-Based Catalyzed Hydroformylation of Short-Chain Olefins, Catalysts , 2020, 10 (5), 510, DOI: 10.3390/catal10050510 .
- K. Ghaib, K. Nitz and F.-Z. Ben-Fares, Chemical Methanation of CO 2 : A Review, ChemBioEng Rev. , 2016, 3 (6), 266–275, DOI: 10.1002/cben.201600022 .
- M. Tkachenko, M. Malyuk, A. Holmanyuk and N. Liubimov, Label Studio: Data labeling software.
- B. Motik, R. Shearer, G. Stoils and I. Horrocks, HermiT OWL Reasoner: The New Kid on the OWL Block, University of Oxford, Accessed: May 14 2022. [Online]. Available: https://www.hermit-reasoner.com/.
- A. S. Behr and D. Chernenko, CatalysisIE Knowledge Graph Generator. [Online]. Available: https://github.com/AleSteB/CatalysisIE_Knowledge_Graph_Generator.
| † Electronic supplementary information (ESI) available: https://github.com/AleSteB/CatalysisIE_Knowledge_Graph_Generator. See DOI: |
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Open access
- Published: 07 August 2024
Male autism spectrum disorder is linked to brain aromatase disruption by prenatal BPA in multimodal investigations and 10HDA ameliorates the related mouse phenotype
- Christos Symeonides ORCID: orcid.org/0009-0009-9415-4097 1 , 2 , 3 na1 ,
- Kristina Vacy ORCID: orcid.org/0009-0000-5330-5260 4 , 5 na1 ,
- Sarah Thomson ORCID: orcid.org/0000-0002-5120-3997 4 ,
- Sam Tanner ORCID: orcid.org/0009-0003-9363-0756 4 ,
- Hui Kheng Chua ORCID: orcid.org/0000-0002-6047-4027 4 , 6 ,
- Shilpi Dixit ORCID: orcid.org/0000-0003-4837-0548 4 ,
- Toby Mansell ORCID: orcid.org/0000-0002-1282-6331 2 , 7 ,
- Martin O’Hely ORCID: orcid.org/0000-0002-0212-1207 2 , 8 ,
- Boris Novakovic ORCID: orcid.org/0000-0002-5623-9008 2 , 8 ,
- Julie B. Herbstman 9 , 10 ,
- Shuang Wang ORCID: orcid.org/0000-0002-1693-6888 9 , 11 ,
- Jia Guo ORCID: orcid.org/0000-0002-9774-9856 9 , 11 ,
- Jessalynn Chia 4 ,
- Nhi Thao Tran ORCID: orcid.org/0000-0002-0396-9760 4 nAff28 ,
- Sang Eun Hwang ORCID: orcid.org/0009-0009-7271-7493 4 ,
- Kara Britt ORCID: orcid.org/0000-0001-6069-7856 12 , 13 , 14 ,
- Feng Chen 4 ,
- Tae Hwan Kim ORCID: orcid.org/0009-0000-6163-3483 4 ,
- Christopher A. Reid 4 ,
- Anthony El-Bitar 4 ,
- Gabriel B. Bernasochi ORCID: orcid.org/0000-0002-3966-2074 4 , 15 ,
- Lea M. Durham Delbridge ORCID: orcid.org/0000-0003-1859-0152 15 ,
- Vincent R. Harley ORCID: orcid.org/0000-0002-2405-1262 12 , 16 ,
- Yann W. Yap 6 , 16 ,
- Deborah Dewey ORCID: orcid.org/0000-0002-1323-5832 17 ,
- Chloe J. Love ORCID: orcid.org/0000-0002-2024-4083 8 , 18 ,
- David Burgner ORCID: orcid.org/0000-0002-8304-4302 2 , 7 , 19 , 20 ,
- Mimi L. K. Tang 2 , 15 ,
- Peter D. Sly ORCID: orcid.org/0000-0001-6305-2201 8 , 21 , 22 ,
- Richard Saffery ORCID: orcid.org/0000-0002-9510-4181 2 ,
- Jochen F. Mueller ORCID: orcid.org/0000-0002-0000-1973 23 ,
- Nicole Rinehart ORCID: orcid.org/0000-0001-6109-3958 24 ,
- Bruce Tonge ORCID: orcid.org/0000-0002-4236-9688 25 ,
- Peter Vuillermin ORCID: orcid.org/0000-0002-6580-0346 2 , 8 , 18 ,
- the BIS Investigator Group ,
- Anne-Louise Ponsonby ORCID: orcid.org/0000-0002-6581-3657 2 , 3 , 4 na2 &
- Wah Chin Boon 4 , 26 na2
Nature Communications volume 15 , Article number: 6367 ( 2024 ) Cite this article
10k Accesses
321 Altmetric
Metrics details
- Autism spectrum disorders
- Epigenetics and behaviour
Male sex, early life chemical exposure and the brain aromatase enzyme have been implicated in autism spectrum disorder (ASD). In the Barwon Infant Study birth cohort ( n = 1074), higher prenatal maternal bisphenol A (BPA) levels are associated with higher ASD symptoms at age 2 and diagnosis at age 9 only in males with low aromatase genetic pathway activity scores. Higher prenatal BPA levels are predictive of higher cord blood methylation across the CYP19A1 brain promoter I.f region ( P = 0.009) and aromatase gene methylation mediates ( P = 0.01) the link between higher prenatal BPA and brain-derived neurotrophic factor methylation, with independent cohort replication. BPA suppressed aromatase expression in vitro and in vivo. Male mice exposed to mid-gestation BPA or with aromatase knockout have ASD-like behaviors with structural and functional brain changes. 10-hydroxy-2-decenoic acid (10HDA), an estrogenic fatty acid alleviated these features and reversed detrimental neurodevelopmental gene expression. Here we demonstrate that prenatal BPA exposure is associated with impaired brain aromatase function and ASD-related behaviors and brain abnormalities in males that may be reversible through postnatal 10HDA intervention.
Similar content being viewed by others

Maternal diabetes-mediated RORA suppression in mice contributes to autism-like offspring through inhibition of aromatase

Sex differences in the effects of prenatal bisphenol A exposure on autism-related genes and their relationships with the hippocampus functions

Long term transcriptional and behavioral effects in mice developmentally exposed to a mixture of endocrine disruptors associated with delayed human neurodevelopment
Introduction.
Autism spectrum disorder (ASD or autism) is a clinically diagnosed neurodevelopmental condition in which an individual has impaired social communication and interaction, as well as restricted, repetitive behavior patterns 1 . The estimated prevalence of ASD is approximately 1–2% in Western countries 2 , with evidence that the incidence of ASD is increasing over time 3 . While increased incidence is partly attributable to greater awareness of ASD 4 , other factors including early life environment, genes and their interplay are important 5 . Strikingly, up to 80% of individuals diagnosed with ASD are male, suggesting sex-specific neurodevelopment underlies this condition 5 .
Brain aromatase, encoded by CYP19A1 and regulated via brain promoter I.f 6 , 7 , 8 converts neural androgens to neural estrogens 9 . During fetal development, aromatase expression within the brain is high in males 10 in the amygdala 11 , 12 . Notably, androgen disruption is implicated in the extreme male brain theory for ASD 13 , and postmortem analysis of male ASD adults show markedly reduced aromatase activity compared to age-matched controls. Furthermore, CYP19A1 aromatase expression was reduced by 38% in the postmortem male ASD prefrontal cortex 14 , as well as by 52% in neuronal cell lines derived from males with ASD 15 . Environmental factors, including exposure to endocrine-disrupting chemicals such as bisphenols, can disrupt brain aromatase function 16 , 17 , 18 .
Early life exposure to endocrine-disrupting chemicals, including bisphenols, has separately been proposed to contribute to the temporal increase in ASD prevalence 19 . Exposure to these manufactured chemicals is now widespread through their presence in plastics and epoxy linings in food and drink containers and other packaging products 20 . Although bisphenol A (BPA) has since been replaced by other bisphenols such as bisphenol S in BPA-free plastics, all bisphenols are endocrine-disrupting chemicals that can alter steroid signaling and metabolism 21 . Elevated maternal prenatal BPA levels are associated with child neurobehavioral issues 20 including ASD-related symptoms 22 , 23 , with many of these studies reporting sex-specific effects 20 , 22 , 23 , 24 . Furthermore, studies in rodents have found that prenatal BPA exposure is associated with gene dysregulation in the male hippocampus accompanied by neuronal and cognitive abnormalities in male but not female animals 20 , 23 , 24 . One potential explanation is that epigenetic programming by bisphenols increases aromatase gene methylation, leading to its reduced cellular expression 16 and a deficiency in aromatase-dependent estrogen signaling. If such is the case, it is possible that estrogen supplementation, such as with 10-hydroxy-2-decenoic acid (10HDA), a major lipid component of the royal jelly of honeybees, may be relevant as a nutritional intervention for ASD. Indeed, 10HDA is known to influence homeostasis through its intracellular effects on estrogen responsive elements that regulate downstream gene expression 25 , 26 , as well as its capacity to influence neurogenesis in vitro 27 .
Here, we have investigated whether higher prenatal BPA exposure leads to an elevated risk of ASD in males and explore aromatase as a potential underlying mechanism. We demonstrate in a preclinical (mouse) model that postnatal administration of 10HDA, an estrogenic fatty acid, can ameliorate ASD-like phenotypes in young mice prenatally exposed to BPA.
Human studies
We examined the interplay between prenatal BPA, aromatase function and sex in relation to human ASD symptoms and diagnosis in the Barwon Infant Study (BIS) birth cohort 28 . By the BIS cohort health review at 7-11 years (mean = 9.05, SD = 0.74; hereafter referred to as occurring at 9 years), 43 children had a pediatrician- or psychiatrist- confirmed diagnosis of ASD against the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5) criteria, as of the 30th of June 2023. ASD diagnosis was over-represented in boys with a 2.1:1 ratio at 9 years (29 boys and 14 girls; Supplementary Table 1 ). In BIS, the DSM-5 oriented autism spectrum problems (ASP) scale of the Child Behavior Checklist (CBCL) at age 2 years 29 predicted diagnosed autism strongly at age 4 and moderately at age 9 in receiver operating characteristic (ROC) curve analyses; area under the curve (AuC) of 0.92 (95% CI 0.82, 1.00) 30 and 0.70 (95% CI 0.60, 0.80), respectively. The median CBCL ASP score in ASD cases and non-cases at 9 years was 51 (IQR = 50, 58) and 50 (IQR = 50, 51), respectively. Only ASD cases with a pediatrician-confirmed diagnosis of ASD against the DSM-5, as verified by the 30th of June 2023, were included in this report. We thus examined both outcomes (ASP scale and ASD diagnosis) as indicators of ASD over the life course from ages 2 to 9 years (Supplementary Table 1 ). Quality control information for the measurement of BPA is presented in Supplementary Table 2 .
BPA effects on ASD symptoms at age 2 years are most evident in boys genetically predisposed to low aromatase enzyme activity
Of the 676 infants with CBCL data in the cohort sample, 249 (36.8%) had an ASP score above the median based on CBCL normative data (Supplementary Table 1 ). From a whole genome SNP array (Supplementary Methods), a CYP19A1 genetic score for aromatase enzyme activity was developed based on five single nucleotide polymorphisms (SNPs; rs12148604, rs4441215, rs11632903, rs752760, rs2445768) associated with lower estrogen levels 31 . Among 595 children with prenatal BPA and CBCL data, those in the top quartile of the genetic predisposition score, that is, children with three or more variants associated with lower levels of estrogens were classified as ‘low aromatase activity’ with the remaining classified as ‘high aromatase activity’ (Fig. 1 ). Regression analyses stratified by this genetic score and child’s sex were performed and an association between high prenatal BPA exposure (top quartile (>2.18 μg/L) and greater ASP scores was only seen in males with low aromatase activity, with a matched OR of 3.56 (95% CI 1.13, 11.22); P = 0.03 (Supplementary Table 4 ). These findings were minimally altered following adjustment for additional potential confounders. Among males with low aromatase activity, the fraction with higher than median ASP scores attributable to high BPA exposure (the population attributable fraction) was 11.9% (95% CI 4.3%, 19.0%). These results indicate a link between low aromatase function and elevated ASP scores. A sensitivity analysis using an independent weighted CYP19A1 genetic score confirmed these findings. For the additional score, the Genotype-Tissue Expression (GTEx) portal was first used to identify the top five expression quantitative trait loci (eQTLs; rs7169770, rs1065778, rs28757202, rs12917091, rs3784307) for CYP19A1 in any tissue type that showed a consistent effect direction in brain tissue. A functional genetic score was then computed for each BIS participant by summing the number of aromatase-promoting alleles they carry across the five eQTLs, weighted by their normalized effect size (NES) in amygdala tissue. This score captures genetic contribution to cross-tissue aromatase activity with a weighting towards the amygdala, a focus in our animal studies. The score was then reversed so that higher values indicate lower aromatase activity and children in the top quartile were classified as ‘low aromatase activity’ with the remaining classified as ‘high aromatase activity’. Again, a positive association between prenatal BPA exposure and ASP scores was only seen in males with low aromatase activity, with a matched OR of 3.74 (95% CI 1.12, 12.50); P = 0.03. Additional adjustment for individual potential confounders provided matched ORs between 3.13 to 3.85 (Supplementary Table 5 ).

Conditional logistic regression models were run where participants were matched on ancestry and time of day of urine collection and, for ASD diagnosis at 9 years, each case within these matched groups was individually matched to eight controls based on nearest date of and age at year 9 interview. BPA was classified in quartiles with the top quartile above 2.18 μg/L as high BPA exposure vs the other three quartiles. ‘Low aromatase enzyme activity’ means being in the top quartile and ‘high aromatase enzyme activity’ means being in the lower three quartiles of an unweighted sum of the following genotypes associated with lower estrogen levels 31 (participant given 1 if genotype is present, 0 if not): CC of rs12148604, GG of rs4441215, CC of rs11632903, CC of rs752760, AA of rs2445768. ‘Greater ASD symptoms’ represents a T-score above 50 (that is, above median based on normative data) on the DSM-5-oriented autism spectrum problems scale of the Child Behavior Checklist for Ages 1.5-5 (CBCL). Data are OR ± 95% CI. Source data are provided as a Source Data file. * Since there were only two ASD cases at age 9 in the girls with low aromatase enzyme activity group, the regression model was not run.
BPA effects on ASD diagnosis at 9 years are most evident in boys genetically predisposed to low aromatase enzyme activity
In subgroup analyses where we stratified by child’s sex and unweighted CYP19A1 genetic score, the results were consistent with those found at 2 years. A positive association between high prenatal BPA exposure and ASD diagnosis was only seen in males with low aromatase activity, with a matched OR of 6.24 (95% CI 1.02, 38.26); P = 0.05 (Supplementary Table 4 ). In this subgroup, the fraction of ASD cases attributable to high BPA exposure (the population attributable fraction) was 12.6% (95% CI 5.8%, 19.0%). In a sensitivity analysis where the weighted CYP19A1 genetic score was used, a similar effect size was observed in this subgroup; matched OR = 6.06 (95% CI 0.93, 39.43), P = 0.06 (Supplementary Table 4 ).
Higher prenatal BPA exposure predicts higher methylation of the CYP19A1 brain promoter PI.f in human cord blood
We investigated the link between BPA and aromatase further by evaluating epigenetic regulation of the aromatase gene at birth in the same BIS cohort. CYP19A1 (in humans; Cyp19a1 in the mouse) has eleven tissue-specific untranslated first exons under the regulation of tissue-specific promoters. The brain-specific promoters are PI.f 6 , 7 , 8 and PII 17 . For a window positioned directly over the primary brain promoter PI.f, higher BPA was positively associated with average methylation, mean increase = 0.05% (95% CI 0.01%, 0.09%); P = 0.009 (Fig. 2 ). Higher BPA levels predicted methylation across both PI.f and PII as a composite, mean increase per log 2 increase = 0.06% (95% CI 0.01%, 0.10%); P = 0.009. Methylation of a control window, comprising the remaining upstream region of the CYP19A1 promoter and excluding both PI.f and PII brain promoters, did not associate with BPA, P = 0.12. These findings persisted after adjustment for the CYP19A1 genetic score for aromatase enzyme activity. Thus, higher prenatal BPA exposure was associated with increased methylation of brain-specific promoters in CYP19A1 . Sex-specific differences were not observed. While these effects were identified in cord blood, methylation of CYP19A1 shows striking concordance between blood and brain tissue (Spearman’s rank correlation across the whole gene: ρ = 0.74 (95% CI 0.59, 0.84); over promoter PI.f window: ρ = 0.94 (95% CI 0.54, 0.99) 32 . Thus, prenatal BPA exposure significantly associates with disruption of the CYP19A1 brain promoter and hence likely the level of its protein product, aromatase.

Visualized using the coMET R package. A Association of individual CpGs along the region of interest with BPA exposure, overlaid with three methylation windows: a 2 CpG window positioned directly on promoter PII, and 7 and 15 CpG windows overlapping PI.f. The red shading reflects each CpG’s level of methylation (beta value). B The CYP19A1 gene, running right to left along chromosome 15, and the positions of both brain promoters. Orange boxes indicate exons. C A correlation matrix for all CpGs in this region. Highlighted in tan are the two CpGs located within the PII promoter sequence and the single CpG located within PI.f. For the 7 CpG window over promoter PI.f, higher BPA associated positively with methylation, mean increase = 0.05% (95% CI 0.01%, 0.09%); P = 0.009, after adjustment for relevant covariates including cell composition. The BPA-associated higher methylation of the brain promoter PI.f region remained evident when the window was expanded to 15 CpGs (mean increase = 0.06%, 95% CI [0.01%, 0.11%], P = 0.04). For PII, the BPA-associated mean methylation increase was 0.07%, 95% CI [-0.02%, 0.16%], P = 0.11). BPA also associated positively with methylation across both PI.f and PII as a composite, mean increase = 0.06% (95% CI 0.01%, 0.10%); P = 0.009. For the remainder of CYP19A1 , excluding both PI.f and PII brain promoters, there was no significant association, P = 0.12. Higher CYP19A1 brain promoter methylation leads to reduced transcription 17 . All statistical tests are two sided. Source data are provided as a Source Data file.
Replication of the association between higher BPA levels and hypermethylation of the CYP19A1 brain promoter
Previously, the Columbia Centre for Children’s Health Study-Mothers and Newborns (CCCEH-MN) cohort (Supplementary Table 3 ) found BPA increased methylation of the BDNF CREB-binding region of promoter IV both in rodent blood and brain tissue at P28 and in infant cord blood in the CCCEH-MN cohort 33 . In rodents, BDNF hypermethylation occurred concomitantly with reduced BDNF expression in the brain 33 . Re-examining the CCCEH-MN cohort, BPA level was also associated with hypermethylation of the aromatase brain promoter P1.f (adjusted mean increase 0.0040, P = 0.0089), replicating the BIS cohort finding.
Molecular mediation of higher BPA levels and hypermethylation of BDNF through higher methylation of CYP19A1
In BIS, we aimed to reproduce these BDNF findings and extend them to investigate aromatase methylation as a potential mediator of the BPA- BDNF relationship. A link between aromatase and methylation of the BDNF CREB-binding region is plausible given that estrogen (produced by aromatase) is known to elevate brain expression of CREB 34 , 35 . In BIS, male infants exposed to BPA (categorized as greater than 4 µg/L vs. rest, following the CCCEH-MN study) had greater methylation of the BDNF CREB-binding site (adjusted mean increase = 0.0027, P = 0.02). This was also evident overall (adjusted mean increase = 0.0023, P = 0.006), but not for females alone (adjusted mean increase = 0.0019, P = 0.13). We then assessed whether methylation of aromatase promoter P1.f mediates this association. In both cohorts, aromatase methylation was positively associated with BDNF CREB-binding-site methylation in males (BIS, adjusted mean increase = 0.07, P = 0.0008; CCCEH-MN, adjusted mean increase = 0.91, P = 0.0016). In the two overall cohorts, there was evidence that the effect of increased BPA on BDNF hypermethylation was mediated partly through higher aromatase methylation (BIS, indirect effect, P = 0.012; CCCEH-MN, indirect effect, P = 0.012).
Prenatal programming laboratory studies—BPA effects on cellular aromatase expression in vitro, neuronal development as well as behavioral phenotype in mice
Bpa reduces aromatase expression in human neuroblastoma sh-sy5y cell cultures.
To validate the findings of our human observational studies on BPA and aromatase expression, we began by studying the effects of BPA exposure on aromatase expression in the human neuroblastoma cell line SH-SY5Y (Fig. 3A ). Indeed, the aromatase protein levels more than halved in the presence of BPA 50 μg/L ( P = 0.01; Fig. 3B ) by Western Blot analysis.

A Western Blot (1 representative blot) demonstrates that increasing BPA concentrations reduced immunoblotted aromatase protein signals (green fluorescence, 55 kDa) in lysates from human-derived neuroblastoma SH-SY5Y cells. Each sample was normalized to its internal house keeping protein β-Actin (red fluorescence, 42 kDa). B Aromatase immunoblotted signals in SH-SY5Y cells treated with vehicle or BPA ( n = 3 independent experiments/group). Five-day BPA treatment of SH-SY5Y cells leads to a significant reduction in aromatase following 50 mg/L(MD = 89, t(6) = 4.0, P = 0.01) and 100 mg/L (MD = 85, t(6) = 3.9, P = 0.01) BPA treatment, compared to vehicle. C BPA treatment (50 µg/kg/day) of Cyp19 -EGFP mice at E10.5-E14.5 results in fewer EGFP+ neurons in the medial amygdala (MD = -5334, t(4) = 5.9, P = 0.004) compared to vehicle mice, n = 3 mice per treatment. Independent t -tests were used and where there were more than two experimental groups ( B ), P -values were corrected for multiple comparisons using Holm-Sidak. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file. Note: UT = untreated.
The effects of prenatal BPA exposure on aromatase-expressing neurons within the amygdala of male mice
There is a prominent expression of aromatase within cells of the male medial amygdala (MeA) 11 . To visualize aromatase-expressing cells, we studied genetically modified, Cyp19 -EGFP transgenic mice harboring a single copy of a bacterial artificial chromosome (BAC) encoding the coding sequence for enhanced green fluorescent protein (EGFP) inserted upstream of the ATG start codon for aromatase ( Cyp19a1 ) 11 (see Methods). As shown, EGFP (EGFP+) expression in male mice was detected as early as embryonic day (E) 11.5 (Supplementary Fig. 1 ), indicating that aromatase gene expression is detectable in early CNS development.
To study the effects of prenatal BPA exposure on brain development, pregnant dams were subject to BPA at a dose of 50 μg/kg/day via subcutaneous injection, or a vehicle injection during a mid-gestation window of E10.5 to E14.5, which coincides with amygdala development. This dose matches current USA recommendations 36 , 37 as well as the Tolerable Daily Intake (TDI) set by the European Food Safety Authority (EFSA) at the time that the mothers in our human cohort were pregnant 28 , 38 . In these experiments, we observed that prenatal BPA exposure led to a 37% reduction ( P = 0.004) in EGFP+ neurons in the MeA of male EGFP+ mice compared to control mice (Fig. 3C ). These results are consistent with our findings in SH-SY5Y cells that indicate that BPA exposure leads to a marked reduction in the cellular expression of aromatase.
Prenatal BPA exposure at mid-gestation influences social approach behavior in male mice
Next, we evaluated post-weaning social approach behavior (postnatal (P) days P21-P24) using a modified three-chamber social interaction test 39 (Fig. 4C ). As shown, male mice with prenatal exposure to BPA were found to spend less time investigating sex- and age-matched stranger mice, when compared with vehicle-treated males (with a mean time ± SEM of 101.2 sec ± 11.47 vs. 177.3 s ± 26.97, P = 0.0004; Fig. 4A ). Such differences were not observed for female mice prenatally exposed to BPA (Fig. 4A ). As a control for these studies, we found that the presence of the EGFP BAC transgene is not relevant to behavioral effects in the test (Supplementary Fig. 2 ), and the proportions of EGFP transgenic mice were not significantly different across BPA-exposed and vehicle-exposed cohorts.

Sociability is the higher proportion of time spent in the stranger interaction zone compared to the empty interaction zone. In the three chamber social interaction test ( A ) BPA-exposed mice ( n = 30, MD = 75 s, t(96) = 3.7 P = 0.0004) spent less time investigating the stranger mouse as compared with male control ( n = 21) mice. B Male ArKO ( n = 8, MD = 43 s, t(30) = 2.3, P = 0.03) mice also spent less time with the stranger compared to male WT littermates ( n = 9). C A schematic of the 3-Chamber Sociability Trial. Created with BioRender.com. D Male BPA-exposed mice ( n = 12, MD = 8.2, U = 11, P = 0.048.) spent more time grooming compared to control ( n = 5) mice. There were no differences between female BPA-exposed ( n = 9) and female control ( n = 6) mice. Independent t -tests were used P -values were corrected for multiple comparisons using Holm-Sidak. For ( C ), a Mann–Whitney U test was used. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file. Note: Veh = vehicle.
To determine if the effects of prenatal BPA exposure were developmentally restricted, we delivered subcutaneous injections (50 µg/kg/day) of BPA to pregnant dams at early (E0.5–E9.5), mid (E10.5–E14.5), and late (E15.5–E20.5) stages of gestation. From these experiments, we found that while male pups exposed to BPA in mid-gestation developed a social approach deficit, such behavioral impairments were not observed for early or late gestation BPA exposure (Supplementary Fig. 3 ). In addition, we performed experiments in which BPA was available to dams by voluntary, oral administration (50 µg/kg/day) during mid-gestation. As shown, a social approach deficit was again observed in male mice (Supplementary Fig. 4 ), consistent with results from prenatal (mid-gestation) BPA exposure by subcutaneous injections. Thus, we find that prenatal BPA exposure at mid-gestation (E10.5-E14.5) in mice leads to reduced social approach behavior in male, but not female offspring. Notably, the amygdala of embryonic mice undergoes significant development during mid-gestation 40 .
Aromatase knockout (ArKO) male mice have reduced social behavior
Having demonstrated that prenatal BPA exposure reduces aromatase expression in SH-SY5Y cells and affects the postnatal behavior of mice, we next asked if the aromatase gene ( Cyp19a1 ) is central to these phenotypes. To address this, we performed social approach behavioral testing (Supplementary Fig. 5 ) on aromatase knockout (ArKO) mice 41 which have undetectable aromatase expression. The social preference towards the stranger interaction zone compared to the empty zone was only evident for the wildtype ( P = 0.003 Fig. 4B ) but not the ArKO ( P = 0.45 Fig. 4B ). This male-specific social interaction deficit is similar to the BPA exposed pups. Further, postnatal estrogen replacement could reverse the ArKO reduction in sociability seen in males ( P = 0.03 Supplementary Fig. 5 ) resulting in a similar stranger-to-empty preference in the E2-treated ArKO as observed for wildtype. The female ArKO pups did not have a sociability deficit (Supplementary Fig. 5 ).
Further, we did not observe any behavioral differences between ArKO vs WT (or BPA exposed vs unexposed) mice of both sexes in Y-maze test. All groups were able to distinguish the novel arm from the familiar arm. All groups spent significantly more time in the novel arm compared to the familiar arm (Supplementary Fig. 6 ), excluding major short-term memory, motor and sensory intergroup difference contributions.
Prenatal exposure to BPA affects repetitive behavior in male mice
Using the water squirt test, we have previously reported that male ArKO, but not female ArKO mice displayed excessive grooming, a form of repetitive behavior, compared to WT mice 42 . Thus, we conducted the water squirt test on BPA-exposed mice to find that male but not female mice exhibited excessive grooming behavior ( P = 0.048; Fig. 4D). Thus, male prenatal BPA-exposed mice and ArKO mice, but not females, exhibited such repetitive behaviors compared to control mice.
The development of the MeA is altered in male ArKO mice as well as in prenatal BPA-exposed male mice
The development and function of the amygdala are highly relevant to human brain development and ASD 43 , 44 . Notably, the medial amygdala (MeA) is central to emotional processing 45 , and this tissue is a significant source of aromatase-expressing neurons. Given that aromatase function in the amygdala is significant for human cognition 46 and behavior 12 , 47 , and that aromatase is highly expressed in the mammalian MeA, as particularly observed in male mice 11 , we investigated changes to the structure and function of this brain region. We performed stereology analyses on cresyl violet (Nissl)-stained sections of male MeA, we observed a 13.5% reduction in neuron (defined by morphology, size, and presence of nucleolus) number. Compared to the vehicle-exposed males, BPA-exposed males had significantly reduced total neuron number (mean count of 91,017 ± SEM of 2728 neurons vs 78,750 ± SEM of 3322 neurons, P = 0.046; Supplementary Fig. 7 ).
We further examined the characteristics of cells within this amygdala structure in detail using Golgi staining (Fig. 5A, B ). We found that the apical and basal dendrites in the MeA were significantly shorter in male BPA-exposed mice vs. vehicle-treated mice (apical: 29.6% reduction, P < 0.0001; basal, P < 0.0001). This phenotype was also observed for male ArKO vs. WT mouse brains (apical 35.0% reduction, P < 0.0001; basal 31.9% reduction, P < 0.0001; Fig. 5A ). Dendritic spine densities of apical and basal dendrites of male ArKO mice, as well as male mice exposed to BPA, were also significantly reduced (KO vs WT apical, P = 0.01; KO vs WT basal, P < 0.0001; BPA treated vs vehicle treated apical P < 0.0001; BPA treated vs vehicle treated basal, P = 0.004; Fig. 5B ). The dendritic lengths (Fig. 5A ) and spine densities (Fig. 5B ) for apical and basal neurites within the MeA of female ArKO mice or BPA-exposed mice were not significantly different compared to control. Thus, in the context of aromatase suppression by prenatal BPA-exposure, or in ArKO mice lacking aromatase, we find that the apical and basal dendrite features within the MeA are affected in a sexually dimorphic manner.

A Golgi staining showed shorter apical and basal dendrites in male BPA-exposed (apical: n = 27β = −136 μ m, 95% CI [−189, −83], P = 6.0 × 10 −7 ; basal: n = 27 neurons β =-106, 95% CI [−147, −64], P = 5.1 × 10 −7 ) and ArKO mice (apical: n = 27 β = −194, 95% CI [−258, −130], P = 2.9 × 10 −9 ; basal: n = 27, β = −121, 95% CI [−143, −100], P = 1.2 × 10 −29 ) compared to male vehicle (apical n = 27, basal n = 27) or WT (apical n = 27, basal n = 29). Female BPA-exposed mice had longer basal dendrites vs. vehicle ( n = 26 neurons/group, β = 133, 95% CI [76, 191], P = 5.5 × 10 −6 ), while female ArKO mice had shorter basal dendrites vs. WT ( n = 22 neurons/group, β = −45, 95% CI [−91, −0.2], P = 0.049). Significant sex-by-BPA-treatment interaction effects were observed for apical ( P = 0.0002) and basal ( P = 3.0 × 10 −11 ) dendritic lengths, and a sex-by-genotype interaction for basal length ( P = 0.003) but not apical length ( P = 0.19). B Golgi staining showed male BPA-exposed (apical: n = 39, β = −7.0, 95% CI [−10.1, −4.0], P = 5.5 × 10 −6 ; basal: n = 90, β = −3.4, 95% CI [−5.7, −1.1], P = 0.004) and ArKO (apical: n = 51, β = −6.8, 95% CI [−12.2, −1.3], P = 0.01; basal: n = 97, β = −3.8, 95% CI [−5.4, −2.2], P = 5.2 × 10 −6 ) mice had lower spine densities on apical and basal dendrites vs. vehicle (apical n = 46, basal n = 106) or WT (apical n = 53, basal n = 109) mice. Female mice exhibited no spine density differences for BPA exposure (apical n = 74, basal n = 106) vs. vehicle (apical n = 61, basal n = 103) and ArKO (apical n = 94, basal n = 86) vs. WT (apical n = 88, basal n = 83). There was a significant sex-by-BPA-treatment interaction for apical spine density ( P = 0.0005) but not basal ( P = 0.99), and no significant sex-by-genotype interactions (apical: P = 0.08; basal: P = 0.19). For golgi staining experiments, 3 mice/group with 6–9 neuron measures/mouse. Spine count datapoints represent the number of spines on a single 10μm concentric circle. C c-Fos fluorescent immunostaining in adult male PD-MeA revealed fewer c-Fos+ve cells in BPA-exposed ( n = 3) vs. vehicle mice ( n = 3; mean difference MD = 3687, t (4) = 16.12, P < 0.0001) and ArKO ( n = 4) vs. WT mice ( n = 4; MD = −10237; t (4) = 6.48, P = 0.0002). Early postnatal estradiol restored ArKO c-Fos to WT levels ( n = 4; MD = −3112; t(4) = 1.97, P = 0.08). D Microelectrode array electrophysiology showed a lower rate of change in EPSP over 1-4 volts in male BPA-exposed mice ( n = 5 mice, n = 11 slices) vs. vehicle ( n = 7 mice, n = 12 slices; P = 0.02). Generalized estimating equations were used clustering by mouse ( A , B ) or voltage input ( D ) and assuming an exchangeable correlation structure. For ( C ), independent t -tests were used and where there were more than two experimental groups (ArKO analysis), P -values were corrected for multiple comparisons using Holm–Sidak. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file.
Prenatal BPA exposure or loss of aromatase in ArKO male mice leads to amygdala hypoactivation and alters behavioral response to a novel social stimulus
The amygdala, a social processing brain region, is hyporesponsive in ASD (see review ref. 48 ). A post-mortem stereology study reported that adolescents and adults diagnosed with ASD feature an ~15% decrease in the numbers of neurons within the amygdala 43 . Also, functional MRI studies report amygdala hypoactivation in participants with ASD compared to controls 49 . Given that the amygdala is a significant source of aromatase-expressing neurons, we next conducted a series of studies to explore how aromatase deficiency influences the male mouse amygdala, using a combination of c-Fos immunohistochemistry, Golgi staining of brain sections, as well as electrophysiological analyses.
To investigate amygdala activation responses after interacting with a stranger mouse, we performed c-Fos immunohistochemistry (a marker for neuronal activation 50 ; Supplementary Fig. 8 ). As shown, prenatal BPA-exposed mice featured 58% fewer c-Fos positive neurons than in the amygdala of vehicle-exposed mice brains ( P < 0.0001; Fig. 5C ). Similarly, we found that the MeA of ArKO mice had a marked deficit of 67% c-Fos-positive neurons when compared with WT ( P = 0.0002) mice, which was ameliorated by early postnatal estradiol replacement (Fig. 5C ). Therefore, prenatal BPA exposure or loss of aromatase expression in ArKO mice leads to amygdala hypoactivation.
Next, we measured the synaptic excitability (I/O curve) of the MeA using multiple electrode analysis, with excitatory postsynaptic potential (EPSP) output indicative of electrical firing by local neurons. As shown, compared to corresponding controls, we find that MeA excitability (I/O curve) is significantly reduced in male mice prenatally exposed to BPA as well as in male ArKO mice (Figs. 5 D and 9D ). As shown, at 4-volt input, BPA treatment resulted in a 22.8% lower ( P = 0.02) excitatory EPSP output than the vehicle treatment, while a 21% reduction ( P = 0.03) in signal was observed for male ArKO mice compared to male WT mice. Thus, prenatal BPA exposure leads to functional hypoactivation of the amygdala of male mice, and this pattern is also evident in male ArKO mice.
Prenatal BPA exposure or loss of aromatase in ArKO male mice leads to abnormalities in neuronal cortical layer V as well as brain function
It has been reported that individuals with ASD show distinct anatomical changes within the somatosensory cortex, including in neurons of cortical layer V 51 . We previously reported that layer V within the somatosensory cortex is disrupted in ArKO mice 52 . Thus, we performed Golgi staining to study the apical and basal dendrites of neurons within layer V of the somatosensory cortex following prenatal BPA exposure, as well as in ArKO mice. As shown, we found that apical and basal dendrite lengths of layer V cortical neurons were significantly decreased in male mice prenatally exposed to BPA, compared with vehicle-treated mice (apical P = 0.04; basal P < 0.0001, Fig. 6A ). Such reductions in dendrites were also reported in male ArKO vs. WT mice (apical P < 0.0001; basal P = 0.02; Fig. 6A ). Furthermore, we found that dendritic spine densities on apical dendrites were also reduced (BPA-exposed mice vs. vehicle, P = 0.04; ArKO vs. WT mice, P = 0.01 (Fig. 6B ).

A Golgi staining showed shorter apical and basal dendrites in male BPA-exposed (apical: n = 36, β =-350μm, 95% CI [−679, −20], P = 0.04; basal: n = 36, β = −217, 95% CI [−315, −119], P = 1.4 × 10 −5 ) and ArKO mice (apical: n = 35, β = −541.9, 95% CI [−666, −417], P = 1.3 × 10 −17 ; basal: n = 35, β = −163, 95% CI [−308, −17], P = 0.02) compared to male vehicle (apical n = 35, basal n = 36) or WT(apical n = 36, basal n = 36). B Golgi staining showed male BPA-exposed (apical: n = 186, β = −4.7, 95% CI [−9.2, −0.2], P = 0.04; basal: n = 40 β = −6.7, 95% CI [−16, 2.8], P = 0.17) and ArKO (apical: n = 148 β = −4.4, 95% CI [−7.7, −1.0], P = 0.01; basal: n = 51 β = −5.2, 95% CI [−14.4, 4.1], P = 0.27) mice had lower spine densities on apical but not on basal dendrites vs. vehicle (apical n = 189, basal n = 56) or WT mice (apical n = 185, basal n = 55). For golgi staining experiments, 3 mice/group with 9–12 neuron measures/mouse. Spine count datapoints represents the number of spines on a single 10 μm concentric circle. C Representative photomicrographs of golgi stained cortical neurons, scale bar is 100μm. D Electrocorticograms (ECoG) revealed an increased in the average spectral power at 8 Hz in BPA-exposed ( n = 4; * a 8 Hz MD = −0.5; t(325) = 3.4 P = 0.01) mice and (E) 4–6 Hz in ArKO mice ( n = 4; * b 4 Hz MD = −0.2; t(120) = 4.3, P = 0.0006; * c 5 Hz MD = −0.2, t (120) = 6.1, P < 0.0001; * d 6 Hz MD = −0.2, t (120) = 5.2 P < 0.0001) vs. vehicle ( n = 7) or WT ( n = 4)mice. Generalized estimating equations were used clustering by mouse (Panels A, B) and assuming an exchangeable correlation structure. For ( D ) and(E) Independent t test were used used, P -values were corrected for multiple comparisons using Holm-Sidak. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file.
To explore the effects of reduced aromatase on cortical activity, we performed electrocorticography (ECoG) recordings from mice in both experimental models (Fig. 6C ). As shown, spectral analysis revealed an increased power in the range of 4–6 Hz for ArKO vs. WT mice (4 Hz P = 0.0006, 5 Hz P < 0.0001, 6 Hz P < 0.0001; Fig. 6D ) and at 8 Hz for BPA-exposed vs. vehicle mice ( P = 0.01; Fig. 6C ). These data indicate that BPA-exposure or loss of aromatase in ArKO mice affects cortical activity, a result which is reminiscent of cortical dysfunction evidenced by EEG recordings on human participants diagnosed with ASD 53 .
Molecular docking simulations indicate 10HDA is acting as a ligand at the same site as BPA on Estrogen Receptors α and β
It has been reported that BPA interferes with estrogen signaling through its competitive interaction and binding with estrogen receptors α (ERα) and β (ERβ) 54 . To explore this in the context of our findings, we used high-resolution in silico 3D molecular docking simulations to model the binding affinity of the natural ligand 17β-estradiol, the putative ligand BPA, as well as a putative therapeutic ligand of interest 10HDA with ERα (Protein Data Bank (PDB) ID: 5KRI) and ERβ (PDB ID: 1YYE). As shown, our spatial analysis indicated that all three ligands have robust binding affinity (Fig. 7 ; Supplementary Movie 1 ). However, while docking alignment revealed that the predicted fit for 10HDA is strikingly similar to that of 17β-estradiol 25 , 55 , BPA showed a greater mismatch (Fig. 7D ), consistent with previous reports that BPA is 1000-fold less estrogenic than the native ligand 56 . Thus, at least for ERα and Erβ, we find that 10HDA may be effective as a competitive ligand that could counteract the effects of BPA on estrogen signaling within cells.

In silico molecular docking analysis of estrogen receptor β (ERβ, Protein Data Bank (PDB) ID: 1YYE; encoded by the ES gene) using the DockThor platform, showing binding predictions for ( A ) the native ligand 17β-estradiol (E2), ( B ) bisphenol A (BPA), ( C ) Trans-10-hydroxy-2-decenoic acid (10HDA), and ( D ) E2 and BPA (left) and E2 and 10HDA (right) superimposed for spatial alignment comparison. While the molecular affinities of BPA and 10HDA for Erβ were comparable (−9.2 vs. −7.9, respectively), 10HDA aligns better with the binding conformation of the endogenous ligand E2, which activates the receptor. BPA is previously reported as sub-optimally estrogenic 106 — >1000-fold less compared to natural estradiol 54 , 56 —whereas 10HDA has an estrogenic role in nature 25 , 55 . Thus, 10HDA may compensate for E2 deficiency caused by a reduction in aromatase enzyme, and in competition with binding by BPA. Please see Supplementary Movie 1 for a video of the above molecular docking of ERβ with E2 superimposed with BPA and Supplementary Movie 2 for the above molecular docking of ERβ with E2 superimposed with 10HDA.
In vitro effects of BPA and 10HDA in primary fetal cortical cell cultures from male brains
Examining male fetal primary cortical culture, BPA alone shortened neurite lengths (Fig. 8A , BPA; quantified in Fig. 8B-C ) and decreased the spine density. BPA treatment reduced both neurite length ( P = 0.0004) and spine densities ( P < 0.0001; Fig. 8A , BPA; quantified in Fig. 8B–C ). Co-administration with 10HDA ameliorated these adverse effects of BPA (Fig. 8A , 10HDA + BPA; quantified in Fig. 8B, C ).

A Representative photomicrographs of primary cultures of embryonic (ED15.5) mouse cortical neurons, red staining is βIII tubulin and green is aromatase. Scale bar is 100μm. B Compared to the BPA group, the vehicle group (β = 79.9, 95% CI [36, 124], P = 0.0004) and BPA + 10HDA group (β = 174, 95% CI [102, 247], P = 2.4 × 10 −7 ) have significantly longer neurites. The BPA + 10HDA group (β = 94, 95% CI [8, 180], P = 0.03) has longer neurites compared to the vehicle group, and there is no difference between the vehicle and 10HDA groups. C Compared to the BPA group, the vehicle group(β = 16, 95% CI [11, 21], P = 1.7 × 10 −8 ) and BPA + 10HDA group (β = 30, 95% CI [21, 40], P = 8.4 × 10 −10 ) have significantly higher spine densities. The BPA + 10HDA group (β = 14, 95% CI [4, 24], P = 0.006) has a higher spine density compared to the vehicle group, and there is no difference between the vehicle and 10HDA groups. n = 10 neurons/group. Primary cortical cell culture was obtained from 12 male mouse embryoes. Spine count datapoints represent the number of spines on a single 10μm concentric circle. Generalized estimating equations were used clustering by mouse and assuming an exchangeable correlation structure. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file.
In vivo effects of 10HDA on BPA mouse model
Guided by our findings in cultured neurons, we next investigated the effects of postnatal 10HDA administration on mice prenatally exposed to BPA at mid-gestation, as follows. After weaning, pups (six litters, 3 weeks of age) were administered daily injections of 10HDA (0 and 500 μg/kg/day; dissolved in saline, i.p.) for 3 weeks, following which pups were assessed for behavioral phenotypes. Strikingly, 10HDA treatment significantly improved social interaction (Fig. 9A ). To determine whether the effect of 10HDA administration is permanent, all treatments were withdrawn for 3 months, and mouse behaviors were subsequently re-tested. Withdrawal of 10HDA treatment in BPA-exposed male mice resulted in a deficit in social interaction (Fig. 9B ), and this deficit was once again ameliorated by a subsequent 10HDA treatment (Fig. 9C ) at 5 months of age, in adulthood. Taken together, these data demonstrate that continuous, postnatal 10HDA administration is effective for ameliorating social interaction deficits in male mice following prenatal BPA exposure.

A 10HDA treatments increased social approach in males ( n = 10/group, MD = 41.14, U = 11, P = 0.03,) but not females ( n = 8/group), compared to saline controls. After 3 months of treatment withdrawal ( B ), male mice (saline n = 8, 10HDA n = 7) no longer spent more time interacting with strangers, compared to vehicle treatment. C When male mice ( n = 8/group) were subsequently treated with a second round of 10HDA, social approach behavior was once again significantly elevated (MD = 39.2, U = 5, P = 0.003), indicative of a rescue of this behavioral effect. D Compared to the WT Saline ( n = 10) group (β = 57.1, 95% CI [47.4, 66.8]), EPSP increases at a 21% lower rate with increasing input in the ArKO Saline ( n = 14) group (β = 45.1 μV, 95% CI [40.1, 50.1], P = 0.03). No differences in slope were detected when comparing the WT Saline group with each of the other two treatment (WT n = 18, KO n = 12) groups. Mann–Whitney U tests were used and for ( D ), Generalized estimating equations were used clustering by voltage input ( D ) and assuming an exchangeable correlation structure. All statistical tests were two-sided. Plots show mean ± SEM. Source data are in a Source Data file. Note: Sal = saline, w/d = withdrawal.
Next, we wanted to determine if hypoactivity arising from the absence of aromatase in the amygdala may be influenced by 10HDA. To address this question, we studied ArKO mice using multiple-electrode analyses, following 3 weeks of treatment with 10HDA (500 μg/kg/day, i.p.). As shown in Fig, 9D , the electrical activity of the male ArKO amygdala treated with 10HDA was similar to male WT activity levels, whereas saline-treated male ArKO amygdala showed significantly lower activity ( P = 0.03) when stimulated by an input/output paradigm, suggesting that 10HDA treatment was effective to compensate the absence of aromatase. Therefore, we interpret these results to suggest that 10HDA restores signaling deficits arising from aromatase deficiency. Given that prenatal BPA exposure suppresses aromatase, 10HDA supplementation may be relevant to aromatase-dependent signaling in that context as well.
Transcriptomic studies of the fetal brain cortex and cortical cell cultures
MiSeq Next-Gen Sequencing was performed on the transcriptome libraries generated from the brain cortex of the E16.5 fetuses after maternal mid-gestation BPA or vehicle exposure. The action of 10HDA was analyzed by RNAseq of transcriptome libraries from total RNA extracted from primary mouse fetal cortical cultures treated with vehicle or 10HDA. Firstly, pathway analysis of the RNAseq data was performed for Gene Ontology (GO) categories using the clusterProfileR R package. No individual pathways in the BPA analysis survived correction for multiple comparisons using an agnostic (non-candidate) approach. Further candidate investigation using the binomial test showed a significant inverse effect of BPA and 10HDA on pathways previously linked to autism 57 , with 10HDA treatment counteracting the effects of BPA on these pathways (Supplementary Fig. 9 ). Based on our Golgi staining experimental findings relating to altered dendrite morphology, we further assessed the category “dendrite extension” as a candidate pathway. Genes in this pathway were downregulated by BPA (Supplementary Figs. 10 A, 9A) and upregulated by 10HDA (Supplementary Figs. 9B , 10B ). More broadly, Fisher’s exact test showed a significant BPA-associated down-regulation ( P = 0.01), and 10HDA-associated up-regulation ( P = 0.0001), of pathways with the terms “axon” and “dendrite”. Notably, the majority (82%; 9 of the top 11 available) mid gestational biological processes whose activity is overrepresented in induced pluripotent stem cells of autism cases vs. controls 57 were impacted by BPA, and in the opposite direction to 10HDA ( P = 0.03; Supplementary Fig. 9 ).
Next, we performed pathway enrichment analysis, also using a candidate pathway approach, of the RNAseq data using Ingenuity (Fig. 10 ). Strikingly, the effects of BPA and 10HDA on gene expression were diametrically opposed across many functional domains (Fig. 10 ). For example, the canonical pathways “Synaptogenesis Signaling pathway” and “CREB signaling” were downregulated by BPA but upregulated by 10HDA. Similarly, key brain functions, e.g., growth of neurites and neural development were down regulated by BPA and reciprocally upregulated by 10HDA (Fig. 10 ). Taken together, prenatal BPA exposure is detrimental to gene expression through a mechanism that may be ameliorated by postnatal 10HDA administration. The full list of differentially expressed genes can be found in Supplementary Dataset 1 .

The Canonical pathways and Disease and Function—Brain pathway databases were selected in Ingenuity. Several key signaling pathways and brain functions were downregulated ( Z -score less than zero) by BPA and also upregulated ( Z -score greater than zero) by 10HDA. 10HDA downregulated four brain disorder related pathways—hyperactive behavior, seizures, seizure disorder and behavioral deficit. Colored boxes indicate significant ( P < 0.05, Fisher’s Exact Test with Benjamani-hochberg) changes in z -score. Boxes shaded in gray indicate non-significant gene expression changes ( P = 0.05 or greater). Source data are provided as a Source Data file.
Here, we report that prenatal BPA exposure leads to ASD endophenotypes in males, and that this involves the actions of the aromatase gene, as well as its functions in brain cells. Our multimodal approach, incorporating both human observational studies and preclinical studies with two mouse models, offer significant insight into how prenatal programming by BPA disrupts aromatase signaling to cause anatomical, neurological, as well as behavioral changes reminiscent of ASD in males.
In the Barwon Infant Study (BIS) human birth cohort the adverse effect of high prenatal BPA exposure on ASD symptoms (ASP score) at age 2, and clinical ASD diagnosis at age 9, was particularly evident among males with a low aromatase enzyme activity genetic score. Further, studying cord blood gene methylation as an outcome we have demonstrated that BPA exposure specifically methylated the offspring CYP19A1 brain promoter in the BIS cohort, replicated in the CCCEH-MN cohort. Previously, in a meta-analysis of two epigenome-wide association studies (EWAS), two CpGs in the region around the brain promoters PI.f and PII were significantly associated ( P < 0.05) with ASD in both EWAS 58 . Past work 16 , and our findings of BPA reduced aromatase expression in a neuronal cell culture and reduced aromatase-eGFP expression in mouse brain, are consistent with the brain-specific suppression of aromatase expression. We also demonstrated that BPA exposure led to a reduction in steady-state levels of aromatase in a neuronal cell line. Further, we replicated past work that higher prenatal BPA levels are associated with BDNF hypermethylation 33 , previously demonstrated to be associated with lower BDNF expression in males 33 .
We find that prenatal BPA exposure at mid-gestation in mice induces ASD-like behaviors in male but not female offspring, concomitant with cellular, anatomical, functional, and behavioral changes (Supplementary Fig. 11 and Supplementary Fig. 12 ). We found that these features were also observed in male ArKO mice, and this is important because aromatase expression is disrupted in BPA-treated male mice. In the Y-maze test, both the ArKO and BPA offspring did not show any differences with the respective control indicating that there are no major memory, sensory or motor issues in these animals. Given the distinct parallels between the effects of BPA that suppresses aromatase, as well as ArKO mice that lack aromatase, we surmise from our studies that BPA disrupts aromatase function to influence the male mouse brain which manifests as: (i) reduced excitatory postsynaptic potentials in the amygdala, (ii) reduced neuron numbers as well as dendritic lengths and spine densities for neurons within the MeA, (iii) altered cortical activity as recorded by ECoG concomitant with decreased dendritic length and spine density in layer IV/V somatosensory cortical neurons, as well as (iv) enhanced repetitive behaviors and reduced social approach to a stranger. These results in mice are consistent with studies with human participants that report abnormal neuronal structure in these comparative regions within the brains of individuals with ASD 51 . Furthermore, we investigated our gene expression dataset and found that the majority of the top biological processes over-represented in cells derived from ASD cases compared to non-cases in a human pluripotent stem cell analysis with a focus on mid gestational brain development 57 were impacted in opposite directions by BPA compared to 10HDA in our gene expression studies on the male mouse brain (Supplementary Fig. 9 ). Of note, the sexually dimorphic effect we report is consistent with work of others demonstrating that prenatal BPA exposure of rodents led to dysregulation of ASD-related genes with neuronal abnormalities, and learning and memory problems only in males 59 .
In our investigations of BPA, we recognized that 10HDA may be a suitable compound as a ligand in the context of brain ER signaling 27 because of its positive effect on gene expression through stimulation of estrogen responsive DNA elements 25 and its role in neurogenesis 27 —characteristics that altogether may compensate for a relative lack of aromatase-generated neural estrogens. Administration of 10HDA alongside BPA protected neuronal cells in culture from the adverse sequelae observed for BPA alone at the same dose. Three weeks of daily postnatal 10HDA treatment significantly enhanced the sociability of the male BPA-exposed mice and dendrite morphology in primary cell culture. The adverse decrease in dendrite lengths and spine densities of the BPA-exposed mice was also corrected by 10HDA administration (Fig. 8 ). Furthermore, postnatal 10HDA treatment restored amygdala electrical activity in the ArKO mice, indicating that 10HDA likely acts downstream of, rather than directly upon, the aromatase enzyme, given that ArKO mice lack functional aromatase. Transcriptomic analyses revealed that 10HDA upregulated, whereas BPA downregulated, gene expression for fetal programming such as for synaptogenesis and growth of neurites. Some of these pathways could be activated by factors downstream of aromatase, such as 17β-estradiol (Supplementary Fig. 13 ). In this study, the ArKO model was useful because it provided an estrogen deficient comparison 41 . We were able to demonstrate that early postnatal E2 administration restored both MeA neural activation and social preference behavior in the ArKO males.
The molecular docking simulations indicate that ERα and ERβ both comprise docking sites for 10HDA and BPA, however, 10HDA is strongly estrogenic 25 , 55 while BPA is greater than 1000-fold less potent than natural estrogen 54 . Such differences in binding are likely relevant to the diverse transcriptomic effects observed in the cells we analyzed by RNAseq.
Strengths of this study include the multimodal approach to test the hypothesis of the interplay of BPA, male sex, and aromatase suppression. In our human epidemiological studies, extensive information was available to allow confounding to be accounted for using matched analyses for the BPA-ASD cohort finding, and findings persisted after adjustment for further individual confounders. Using a modern causal inference technique, molecular mediation 60 we demonstrate in both birth cohorts that aromatase gene promoter I.f methylation underlies the known effect of higher prenatal BPA on BDNF hypermethylation. Other key features that support an underlying causal relationship include: the consistency of the findings across studies in this program (Supplementary Fig. 11 ); and the consistency with which our experimental laboratory work maps to prior studies of people with ASD (summarized in Supplementary Fig. 12 ) in relation to neuronal and structural abnormality in the amygdala 43 and abnormality in amygdala connectivity 44 , and resting-state cortical EEG 53 . Our findings are also consistent with past work indicating reduced prefrontal aromatase levels in individuals with ASD at postmortem 14 , 15 . The finding that BPA-associated gene methylation patterns in the BIS cohort were not sex-specific but that BPA-associated ASD symptoms and clinical diagnosis were more evident in males with a low genetic aromatase score would be consistent with the male vulnerability to BPA reflecting not differential epigenetic programming, but a greater vulnerability to reduced aromatase function in the developing male brain. This is reinforced by the ArKO model which resulted in an ASD-like phenotype in males not females. We have provided experimental evidence not only on the adverse neurodevelopment effects of BPA, but also experimental evidence of the alleviation of the behavioral, neurophysiological, and neuroanatomical defects following postnatal treatment with 10HDA. A human randomized controlled prevention trial that achieved bisphenol A elimination during pregnancy, with a resultant reduction in ASD among male offspring, would be a useful next step to provide further causal evidence of BPA risks but the feasibility and ethics of such an undertaking would be considerable. We demonstrate that postnatal administration of 10HDA may be a potential therapeutic agent that counteracts the detrimental impacts on distinct gene expression signatures directly impacted by prenatal BPA exposure. Furthermore, 10HDA may ameliorate deficits in ArKO mice which further suggests its utility as a replacement therapy for aromatase deficiency.
Two limitations of our human study were that BPA exposure was measured in only one maternal urine sample at 36 weeks, and that the assay may have low sensitivity 61 . We partially redressed the latter by focusing on categorical BPA values, as recommended 61 , and undertook a matched ASD analysis where determinants for BPA variation, such as the urine collection time of day, were matched to reduce misclassification. Also, functional gene expression studies were unavailable for human samples in our study, but whilst the misclassification introduced by a reliance on a SNP based score would likely lead to an underestimation, an effect was still found among males with a low genetic aromatase score. It would be useful in further studies to consider altered aromatase function with a combined epigenetic-genetic score to reflect environment-by-epigenetic and genetic determinants of low aromatase function. Direct brain EWAS measures were not available, but for the key brain promoter PI.f region of CYP19A1 , the brain-blood correlation is very high: Spearman’s rho= 0.94, 95% CI [0.80, 0.98] 32 . Although ASD symptoms (ASP score) at 2 years were based on parent report, we have previously reported that a higher ASP score was predictive of later ASD diagnosis by age 4 30 . ASD diagnosis at age 9 was verified to meet DSM-5 criteria by pediatrician audit of medical records, thereby reducing diagnostic misclassification.
In our preclinical studies, we performed the 10HDA studies and some of the BPA mechanistic studies only on male animals because our extensive laboratory and human studies, with more than 25 analyses, demonstrated that BPA exposure had significantly more adverse effects in males than females. In addition, we performed the RNASeq on the cortex of fetal mice exposed to BPA in vivo, whereas 10HDA was performed in vitro on cortical primary cell culture. This would likely increase the variability between the BPA RNASeq and the 10HDA RNASeq, yet many of the same pathways were impacted but in opposing directions. Furthermore, the changes we observed in our RNAseq data could be due to changes in the cell type or cell state. This could be clarified by future single cell RNAseq experiments now that this specific issue has been identified.
The BPA exposure of mouse dams under our experimental conditions (50 µg/kg bodyweight) matches the current Oral Reference Dose set by the United States Environmental Protection Agencyc, the current safe level set by the U.S. Food and Drug Administration (FDA) 37 , as well as the Tolerable Daily Intake set by the European Food Safety Authority 38 at the time that the mothers in our human cohort were pregnant 28 . The EFSA set a new temporary TDI of 4 µg/kg bodyweight in 2015 62 and, in December 2021, recommended further reducing this by five orders of magnitude to 0.04 ng/kg 63 ; although this was subsequently revised to 0.2 ng/kg in EFSA's scientific opinion published in 2023 64 . Therefore, the timing of this new evidence is particularly pertinent and provides direct human data to support the reduced TDI.
Consistent with typical human exposure in other settings 20 , BPA exposure in our birth cohort was substantially lower than the above, and yet we see adverse effects. Assuming fractional excretion of 1 65 and average daily urine output of 1.6 L 65 , the median urinary bisphenol concentration of 0.68 µg/L—for which we see increased odds of ASD diagnosis—equates to a total daily intake of just 13 ng/kg, given a mean maternal bodyweight of 80.1 kg at time of urine collection. Notably, while we find an adverse association at 13 ng/kg, we do not have sufficient participants with lower exposure to evaluate a safe lower limit of exposure below this. Our findings in cell culture, with concentration 5 µg/L, parallels these human findings in terms of dose response. Although there are limitations in translating concentrations across body compartments without a stronger understanding of pharmacokinetics of BPA, 5 µg/L corresponds to the 90th and 95th percentile of BPA in urine in our human cohort, and allowing for a standard factor of 10 for variability in human sensitivity used when setting TDIs 38 indicates relevance down to at least 0.5 µg/L, below the median urine concentration. Our findings in laboratory animal studies, with exposure of 50 µg/kg bodyweight, are a little higher, as they were designed to correspond to the then current recommendations 36 , 37 , 38 , but implications nevertheless have relevance within the range of exposure in our cohort. Allowing for standard factors of 10 for interspecies variability 38 and variability in human sensitivity 38 , our animal study findings support a TDI at 500 ng/kg or below, which corresponds to the upper 0.5% of our human cohort. The findings of the human study, also allowing for a factor of 10 for variability in human sensitivity 38 , therefore, support a TDI at or below 1.3 ng/kg.
Despite bans on its use in all infant products by the European Union in 2011 and the U.S. FDA in 2012, BPA remains widespread in the environment 66 . The main source of human exposure to BPA is dietary contamination 68 . Bisphenols are used in the production of common food contact materials, and migrate from those materials during use 69 , including polycarbonate food and beverage containers and the epoxy linings of metal food cans, jar lids, and residential drinking water storage tanks and supply systems 64 . Additional sources of exposure include BPA-based dental composites and sealant epoxies, as well as thermal receipts 64 . BPA levels in pregnant women have previously been reported to be higher for young mothers, smokers, lower education, and lower income 70 . A substantial proportion of ASD cases might be prevented at the population level if these findings were causal and prenatal maternal BPA exposure were reduced. Here, exposures in the top quartile of BPA (>2.18 ug/L) correspond to a population attributable fraction (PAF) for males with low aromatase of 12.6% (95% CI 5.8%, 19.0%) although this estimate is imprecise as it is based on low case numbers. The only other available study with data on BPA exposure (>50 ug/L) and ASD provides an estimate in all children of 10.4% 67 . These studies have misclassification issues (e.g., a single urine measure for BPA and, in the Stein et al. study, an ASD diagnosis derived from health care sources 67 ) but these misclassifications are likely non-differential and thus would bias findings towards the null. Additionally, we need to consider that the above findings of RfD/TDI and PAF are based on BPA alone. Factoring in that most exposures occur as part of a chemical mixture adds additional concern 71 . For example prenatal valproic acid exposed mice (an established ASD mouse model) also have a lower brain aromatase expression 72 .
In summary, this multimodal program of work has shown an adverse effect of higher maternal prenatal BPA on the risk of male offspring ASD by a molecular pathway of reduced aromatase function, which plays a key role in sex-specific early brain development. Overall, these findings add to the growing evidence base of adverse neurodevelopmental effects from bisphenol and other manufactured chemical exposure during pregnancy. The case is compelling and supports broader evidence on the need to further reduce BPA exposure, especially in pregnancy. We also envision that our findings will contribute to new interventions for the prevention and/or amelioration of ASD targeting this specific pathophysiological pathway and we have identified one possible neuroprotective agent—10HDA—that has strong laboratory support. This agent now warrants further study, including human safety and efficacy evaluation.
The human Barwon Infant Study cohort study was approved by the Barwon Health Human Research Ethics Committee, and families provided written informed consent. Parents or guardians provided written informed consent at prenatal recruitment and again when the child was 2 years of age. The human Columbia Center for Children’s Environmental Health Mothers and Newborn cohort study was approved by the Institutional Review Boards of Columbia University and the Centers for Disease Control and Prevention, and all participants in the study provided informed consent. All procedures involving mice were approved by the Florey Institute of Neuroscience and Mental Health animal ethics committee and conformed to the Australian National Health and Medical Research Council code of practice for the care and use of animals for scientific purposes and All experiments were designed to minimize the number of animals used, as well as pain and discomfort. This work adheres to the ARRIVE essential 10 guidelines.
The Barwon Infant Study birth cohort
Participants.
From June 2010 to June 2013, a birth cohort of 1074 mother–infant pairs (10 sets of twins) were recruited using an unselected antenatal sampling frame in the Barwon region of Victoria, Australia 28 . Eligibility criteria, population characteristics, and measurement details have been provided previously 28 ; 847 children had prenatal bisphenol A measures available (Supplementary Table 1 ).
Bisphenol A measurement
We used a direct injection liquid chromatography tandem mass spectrometry (LC-MS/MS) method, as previously described in detail 73 . In summary, a 50 µL aliquot of urine was diluted in milli-Q water and combined with isotopically-labeled standards and b-glucuronidase (from E. Coli -K12). Samples were incubated for 90 min at 37 °C to allow for enzymatic hydrolysis of bisphenol conjugates before quenching the reaction with 0.5% formic acid. Samples were centrifuged before analysis, which was performed using a Sciex 6500 + QTRAP in negative electrospray ionization mode. The BPA distribution and quality control attributes for the application of this method to the Barwon Infant Study (BIS) cohort are shown in Supplementary Table 2 .
Child neurodevelopment
Between the ages of seven and ten, a health screen phone call was conducted to gather information on autism spectrum disorder (ASD) diagnoses and symptomology. Out of the 868 individuals who responded to the health screen, 80 had an ASD diagnosis reported by their parents/guardians or were identified as potentially having ASD. The parent-reported diagnoses were confirmed by pediatric audit of the medical documentation to verify an ASD diagnosis as per DSM-5 guidelines. Participants that had a parent-reported diagnosis and then a verified pediatrician diagnosis by 30 June 2023 and whose diagnoses occurred before the date of their 9-year health screen were included as ASD cases in this study’s analyses ( n = 43). Participants were excluded if (i) their parent/guardian responded with ‘Yes’ or ‘Under Investigation’ to the question of an ASD diagnosis on the year-9 health screen but their diagnosis was not verified by 30 June 2023 ( n = 26), or (ii) they had a verified diagnosis of ASD by 30 June 2023 but their date of diagnosis did not precede the date of their year-9 health screen ( n = 15). The DSM-5-oriented autism spectrum problems (ASP) scale of the Child Behavior Checklist for Ages 1.5-5 (CBCL) administered at 2-3 years was also used as an indicator of autism spectrum disorder.
Whole genome SNP arrays
Blood from the umbilical cord was gathered at birth and then transferred into serum coagulation tubes (BD Vacutainer). Following this, the serum was separated using centrifugation as described elsewhere 74 . Genomic DNA was extracted from whole cord blood using the QIAamp DNA QIAcube HT Kit (QIAGEN, Hilden, Germany), following manufacturer’s instructions. Genotypes were measured by Erasmus MC University Medical Center using the Infinium Global Screening Array-24 v1.0 BeadChip (Illumina, San Diego, CA, USA). The Sanger Imputation Service (Wellcome Sanger Institute, Hinxton, UK) was used for imputing SNPs not captured in the initial genotyping using the EAGLE2 + PBWT phasing and imputation pipeline with the Haplotype Reference Consortium reference panel 75 . Detailed methods are provided elsewhere 76 .
Genome-wide DNA methylation arrays and analysis methods can be found in the Supplementary Methods.
Center for Children’s Environmental Health (CCCEH) epigenetic investigations
The study participants consisted of mothers and their children who were part of the prospective cohort at the Columbia Center for Children’s Environmental Health Mothers and Newborn (CCCEH-MN) in New York City (NYC). They were enrolled between the years 1998 and 2003, during which they were pregnant. The age range for these women was between 18 and 35, and they had no prior history of diabetes, hypertension, or HIV. Furthermore, they had not used tobacco or illicit drugs and had initiated prenatal care by the 20th week of their pregnancy. Every participant gave informed consent, and the research received approval from the Institutional Review Boards at Columbia University as well as the Centers for Disease Control and Prevention (CDC) 33 .
Epigenetic methods have been previously described 77 . Briefly, DNA methylation was measured in 432 cord blood samples from the CCCEH-MN cohort using the 450 K array (485,577 CpG sites) and in 264 MN cord blood samples using the EPIC array (866,895 CpG sites) (Illumina, Inc., San Diego, CA, USA).
BPA measures in the CCCEH were based on spot urine samples collected from the mother during pregnancy (range, 24–40 weeks of gestation; mean, 34.0 weeks) 33 , 78 .
Other statistical analysis
Maternal urinary BPA concentrations were corrected for specific gravity to control for differences in urine dilution. Given a high proportion of the sample (46%) had BPA concentrations that were not detected or below the limit of detection (LOD), a dichotomous BPA exposure variable was formed using the 75th percentile as the cut-point. Dichotomizing the measurements in this way is also likely to give similar results regardless of whether indirect or direct analytical methods were used 79 . This is desirable since indirect methods might be flawed and underestimate human exposure to BPA 61 .
To evaluate whether autism spectrum problems at 2 years could be used as a proxy for later ASD diagnosis, receiver operating characteristic (ROC) curve analyses were used. CBCL ASP at age 2 years predicted diagnosed autism strongly at age 4 and moderately at age 9 with an area under the curve of 0.92 (95% CI 0.82, 1.00) and 0.70 (95% CI 0.60, 0.80), respectively.
According to the normative data of the CBCL, T-scores greater than 50 are above the median. Due to a skewed distribution, ASP measurements were dichotomized using this cut point, which has respective positive and negative likelihood ratios of 2.68 and 0.00 in the prediction of verified ASD diagnosis at 4 years and 1.99 and 0.49 in the prediction of verified ASD diagnosis at 9 years.
A CYP19A1 genetic score for aromatase enzyme activity was developed based on five genotypes of single nucleotide polymorphisms (CC of rs12148604, GG of rs4441215, CC of rs11632903, CC of rs752760, AA of rs2445768) that are associated with sex hormone levels 31 . Participants were classified as ‘low activity’ if they were in the top quartile, that is, they had three or more genotypes associated with lower levels of estrogen and as ‘high activity’ otherwise. Conditional logistic regression model analyses investigating the association between prenatal BPA levels and (i) early childhood ASP scores and (ii) verified ASD diagnosis at 9 years were conducted in the full sample, repeated after stratification by child’s sex (assigned at birth based on visible external anatomy), and repeated again after further stratifying by the CYP19A1 genetic score. Matching variables included child’s sex (in the full sample analysis only), ancestry (all four grandparents are Caucasian vs not) and time of day of maternal urine collection (after 2 pm vs before). Within these matched groups, we additionally matched age-9 ASD cases and non-cases based on the date of the health screen and child’s age at the health screen using the following procedure. Each case was matched to a single non-case based on nearest date of and age at health screen. Once all cases had one matched non-case, a second matched non-case was allocated to each case, and so on until all cases had 8 matched non-cases (8 was the most possible in the boys with high aromatase activity sub-sample and so this number was used across all sub-samples). The order by which cases were matched was randomly determined at the start of each cycle.
The guidelines for credible subgroup investigations were followed 80 . Only two categorical subgroup analyses were conducted, and these were informed a priori by previous literature and by initial mouse study findings. The adverse BPA effects in males with low aromatase enzyme activity (as inferred from the CYP19A1 genetic score) were expected to be of higher magnitude, based on the prior probabilities from the laboratory work. A systematic approach was used to evaluate non-causal explanations and build evidence for causal inference, considering pertinent issues such as laboratory artefacts that are common in biomarker and molecular studies 81 .
A second CYP19A1 genetic score was developed for use in sensitivity analyses. The Genotype-Tissue Expression (GTEx) portal was used to identify the top five expression quantitative trait loci (eQTLs) for aromatase in any tissue type that showed a consistent effect direction in brain tissue. A functional genetic score was then computed for each BIS participant by summing the number of aromatase-promoting alleles they carry across the five eQTLs (AA of rs7169770, CC of rs1065778, AA of rs28757202, CC of rs12917091, AA of rs3784307), weighted by their normalized effect size (NES) in amygdala tissue. The score was then reversed so that higher values indicate lower aromatase activity. The score thus captures genetic contribution to reduced cross-tissue aromatase activity with a weighting towards the amygdala, a focus in our animal studies. The variable was dichotomized using the 75th percentile as the cut-point and the above stratified analyses were repeated with this new weighted score replacing the original, unweighted score.
For the human epigenetic investigations, we used multiple linear regression and mediation 60 approaches. As in past work 33 , BPA was classified as greater than 4 μg/L vs less than 1 μg/L in the CCCEH-MN cohort. A comparable classification was used for the BIS cohort, with greater than 4 μg/L vs the rest. In both cohorts the regression and mediation analyses were also adjusted for sex, gestational age, self-reported ethnicity, and cord blood cell proportions. In the BIS cohort, ethnicity was defined as all four grandparents are Caucasian vs not (see Table S3 ). For the CCCEH-MN cohort, ethnicity was defined as Dominican vs African American 33 . We used statistical software packages R v3.6.3 82 and Stata 15.1 83 .
LABORATORY STUDIES
Shsy-5y cell culture study, bpa treatment on aromatase expression in cell culture.
Human neuroblastoma SHSY-5Y cells were chosen because they were known to express aromatase and SH-SY5Y have been used in ASD research 84 . SHSY-5Y cells (CRL-2266, American Type Culture Collection, Virginia, USA) were maintained in Dulbecco’s Modified Eagle Medium (DMEM) (10313-021, Gibco-life technologies, New York (NY), USA) supplemented with 10% heat-inactivated Fetal Bovine Serum (FBS) (12003C-500 mL, SAFC Biosciences, Kansas, USA), 1% penicillin streptomycin (pen/strep) (15140-122, Gibco-life technologies, NY, USA) and 1% L-Glutamine (Q) (25030-081, Gibco-life technologies, NY, USA) at 37 °C in a humidified atmosphere of 95% air and 5% CO 2 . SHSY-5Y cells were grown in 175 cm 2 cell culture flasks (T-175) (353112, BD Falcon, Pennsylvania, USA). Cells were passaged when the seeding density of the T-175 flasks was reached (roughly 80-90% confluence). Cells were passaged by aspirating media from flasks and flasks were then washed once with 10 mL of DPBS (14190-136 Gibo-life technologies, NY, USA) to remove the FBS (inhibits the actions of trypsin). Next, cells were incubated with trypsin (2 mL/T-175 flask) at 37 °C for 5 min to detach cells from the flask wall. To prevent further action of trypsin, media (8 mL/T-175 flask) was added, and contents were pipetted up and down to disperse cell clumps. The cell suspension was then transferred to a 15 mL centrifuge tube (430791, Corning CentriStar, Massachusetts, USA) and centrifuge (CT15RT, Techcomp, Shanghai, China) for 5 min at 1000 RPM at room temperature (RT). The media was then aspirated from these tubes and the cell pellet resuspended in 1 mL of media. Cell viability counts were performed using a hemocytometer (Hausser Scientific, Pennsylvania, USA) to determine the number of live versus dead cells in solution. Two μL of cell suspension was diluted with media (98 μL) and then trypan blue (100 μL) (T8154, Sigma-Aldrich Co., St. Louis, MO, USA) (which labeled dead cells) in a sterile microcentrifuge tube (MCT-175-C-S, Axygen, California, USA). Ten μL of this solution was loaded into the hemocytometer and imaged using a light microscope (DMIL LED, Leica, Germany). Dead cells appeared blue under the microscope because these cells take up the dye whereas live cells were clear (i.e., unstained). Cells were counted in the outer four squares located in each chamber (two chambers, eight squares), with their dimensions known. The average of the eight counts was multiplied by the dilution factor and by 104, yielding the concentration of cells/mL solution. Average cell counts were plotted against treatment groups using GraphPad Prism 9.0 (GraphPad Software, Inc., San Diego, CA).
Bisphenol A (BPA) (239658-50 G, Sigma-Aldrich Co., St. Louis, MO, USA) was used for cell treatment. Prior to treatment, stock solutions of each drug were prepared as stated below. BPA was dissolved in pure ethanol (EA043-2.5 L, Chem supply, South Australia, Australia) and the final concentration of the stock solutions was 0.0435 g/mL. Cells in T-175 flasks were randomly assigned to receive treatment with BPA at a dosage of 100 μg/L, 50 μg/L, 25 μg/L or 0 μg/L (vehicle). There was also a no treatment (no vehicle added) flask.
Cell treatment, protein assay, SDS-Page, and western blotting methods can be found in the Supplementary Methods.
Animal studies
Two colonies of mice, maintained at the Florey Institute, were used in this study. The Aromatase knockout (ArKO) mouse model and the Aromatase-enhanced fluorescent green protein (Cyp19-EGFP) transgenic mouse model. Animals were monitored daily except for weekends. If animals showed general clinical signs, an animal technician or a vet was consulted for advice and euthanasia performed as required.
Mice were maintained under specific pathogen-free (SPF) conditions on a 12 h day/night cycle, with ad libitum water and soybean-free food (catalog number SF06-053, Glen Forrest Stockfeeders, Glen Forrest, Western Australia, Australia). Facial tissues were provided for nesting material, and no other environmental enrichment was provided. The room temperature ranged from 18 °C-23 °C and the humidity ranged from 45%-55%.
The sex of mice was determined by SRY genotyping if fetal, otherwise sex was determined by examining the anogenital region around PND9 and again at weaning. Sex was confirmed by inspecting the gonads during dissection. Cyp19 -EGFP mice were toe and tail clipped for identification and genotyping at PND9, ArKO mice were ear notched and tailed clipped at two weeks of age. The oligonucleotide sequences (custom oligos, Geneworks, Australia) for ArKO, GFP and SRY genotyping can be found in Supplementary Data File 2 .
Aromatase knockout (ArKO) mouse model
The ArKO mouse is a transgenic model having a disruption of the Cyp19a1 gene. Exon IX of the Cyp19a1 gene was replaced with a neomycin-resistant cassette 41 . Homozygous Knockout (KO) and wild-type (WT) offspring were bred by mating heterozygous (het) ArKO parents and then PCR genotyped. ArKO mice were backcrossed onto a C57BL/6 J background strain, >10 generations (obtained from Animal Resources Centre, Western Australia) and the colony maintained at the Florey institute.
Aromatase-enhanced fluorescent green protein ( Cyp19 -EGFP) transgenic mouse model
The Cyp19-EGFP mouse model (backcrossed onto the FVBN background strain >10 generations, obtained from Animal Resources Centre, Western Australia) is a transgenic model having a bacterial artificial chromosome containing the full length of the Cyp19a1 gene with an Enhanced Green Fluorescent Protein (EGFP) gene inserted upstream of the ATG start codon 11 . Thus, EGFP expression is an endogenous marker for Cyp19a1 expression. This allows for the visualization and subsequent localization of EGFP as the marker for aromatase without the use of potentially nonspecific aromatase antibodies 11 . We have previously characterized this transgenic model and its brain expression of EGFP 11 . Based on our characterization studies, this transgenic model does not have phenotypes that are significantly different to wildtype mice.
Early postnatal 17β-estrodiol treatment
Mice were allocated into three groups: (1) WT mice receiving a sham implantation; (2) ArKO mice receiving a sham implantation; and (3) ArKO mice undergoing implantation with a 17β-estradiol pellet (sourced from Innovative Research America). This estradiol pellet was designed to release 0.2 mg of 17β-estradiol steadily over a period of 6 weeks. A corresponding sham pellet, identical in size but devoid of E2, was implanted in the control groups.
The implantation procedure was carried out on postnatal day 5. For anesthesia, mice were exposed to 2% isoflurane (IsoFlo, Abbott Laboratories, VIC, Australia) within an induction chamber. The efficacy of anesthesia was confirmed by the lack of response to foot-pinch stimuli. During the surgical procedure, mice were maintained on a heated pad to regulate body temperature. A small, 5 mm incision was made in the dorsal region for the subcutaneous insertion of the pellet, preceded by an injection of Bupivacane in the same area. Following the implantation, the incision was carefully sutured. Post-surgery, mice were placed in a thermal cage (Therma-cage, Manchester, UK) for recovery and monitoring until they regained consciousness and could be returned to their respective litters. Any mice exhibiting complications such as opened stitches were excluded from the study.
BPA injection administration treatment
Plugged FVBN dams were randomly assigned, blocking by weight gain at E9.5 and litter/cage where applicable, to receive daily scruff subcutaneous injections (24 G x 1”, Terumo, Somerset, New Jersey, USA) of BPA (239658-50 G, Sigma-Aldrich Co., St Louis, MO, USA) in ethanol and peanut oil (Coles, Victoria, Australia), either between E0.5-E9.5, E10.5-E14.5 or E15.5-birth at a dosage of 50 μg/kg (deemed as the safe consumption level by the Food and Drug Administration, FDA) 37 or 0 μg/kg (vehicle) of maternal body weight. The injection volume was 1.68 μL/g bodyweight. Mice were weighed directly before each injection. BPA and vehicle exposed litters did not differ in litter size (Supplementary Fig. 14 ).
10HDA injection administration treatment
Cyp19 -EGFP or ArKO mice were randomly assigned by blocking on sex and litter to receive daily intraperitoneal injections (31 G x 1”, Terumo, Somerset, New Jersey, USA) of 500 μg/kg 10HDA (Matreya, USA) in saline or vehicle saline for 21 consecutive days. The injection volume was 2.1 μL/g bodyweight. Mice were weighed directly before each injection.
BPA oral administration treatment
Plugged FVBN dams were exposed to jelly at E9.5. The jelly contained 7.5% Cottee’s Raspberry Cordial (Coles, Victoria, Australia) and 1% bacteriological agar (Oxoid, Australia) in milli-Q water. The pH was increased to between 6.5-7.5 with a pallet of NaOH to allow the jelly to set. Dams were then randomly assigned, blocking by weight gain at E9.5 and litter/cage where applicable, to receive a daily dose of jelly, which contained either ethanol or BPA dissolved in ethanol, at a dosage of 50 μg/kg or 0 μg/kg (vehicle) of maternal body weight. Dams received doses between E10.5-E14.5, and only dams that were observed to have consumed all the jelly each day were included in the study.
Behavioral paradigms
Three-chamber social interaction test.
The three-chamber social interaction test is extensively used to investigate juvenile and adult social interaction deficits, including in sociability 85 , 86 . BPA exposed pups were habituated in the experimental room on P21, directly after weaning. Following a two-to-three-day habituation, testing was conducted from P24 to P27, as only a maximum of ten mice could be tested during the light phase per day. ArKO mice treated with estrogen or sham pallet began habituation at PND28-29, with testing at PND31-33. Both male and female mice were tested. Testing was performed in a dedicated room for mouse behavior studies; no other animals were present in the room at the time of acclimatization and testing. The temperature of the room was maintained at approximately 21 °C.
The test apparatus, a three-chambered clear plexiglass, measuring 42 cm x 39 cm x 11 cm, had two partitions creating a left, right (blue zones), and center chamber (green zone) in which mice could freely roam via two 4 cm x 5 cm openings in the partitions (Supplementary Fig. 15 ). The two side chambers contained two empty wire cages. A 1 cm wide zone in front of each wire cage was defined as the interaction zone (yellow zones). The chamber was set on a black table for white mice, and on a white covering for black mice to aide tracking.
Each test consisted of two consecutive 10-min trials, a habituation trial (T1) and a sociability trial (T2). T1 allowed the test mouse to habituate, and any bias for either empty interaction zone was noted. For T2, a C57BL/6 J novel stranger mouse matched with the test mouse for age and gender was introduced into the cage on the opposite side to which the test mouse demonstrated an interaction zone bias. Thus, any evidence of sociability is bolstered as interaction zone bias would have to be overcome.
For each trial, the test mouse began in the center chamber, and its activity, both body center point, and nose point was tracked and quantified by TopScan Lite (Clever Sys Inc., Reston VA, USA). In this study, the key measure extracted was the average duration of the nose point in each interaction zone.
Social approach and sociability were analyzed. We define social approach as the time the test mouse’s nose point was tracked in the stranger cage interaction zone. Sociability is the higher proportion of time the test mouse to spends with the nose point in the stranger cage interaction zone compared to the empty cage interaction zone.
Details on the Y-maze and grooming methods can be found in the Supplementary Methods.
Golgi staining
Mice had not undergone any behavioral testing. For Golgi staining and analysis, Wild Type (WT) and Knockout (ArKO) and Cyp19 -EGFP littermate males (aged P65-P70); one mouse from n = 3 litters for each genotype) were deeply anesthetized with isopentane rapidly decapitated and fresh whole brain tissues were collected. Brains were first washed with milli-Q water to remove excess blood and then directly placed in the solution obtained from the FD Rapid GolgiStain TM Kit (FD Neuro-Technologies, Inc., MD, USA). Brains were stored at room temperature in the dark and the solutions were replaced after 24 hours, and the tissues were kept in the solution for two weeks. After two weeks, tissues were transferred into solution C for a minimum of 48 hours at room temperature. For sectioning, brains were frozen rapidly by dipping into isopentane pre-cooled with dry ice, and 100 µm thick coronal sections were cut at -22 °C and mounted on 1% gelatin-coated slides. The sections were then air dried in the dark at room temperature. When sections were completely dry, slides were further processed and rinsed with distilled water and placed in the solution provided in the kit for 10 min and washed again with distilled water followed by dehydration for 5 min each in 50%, 75%, 95%, and 100 % ethanol. Sections were further processed in xylene and mounted with Permount.
Neuron Tracing
Neuron tracing was conducted on the amygdala and somatosensory cortex of BPA-exposed mice (exposed ED10.5-14.4) and untreated ArKO mice. Neuron tracing in the amygdala was conducted in both male and female mice, and in the somatosensory cortex, only in male mice. Stained slides were coded to ensure that morphological analysis was conducted by an observer who was blind to the animals’ treatment. Morphological analysis followed a previously described protocol 87 with the following modifications: layer V pyramidal cells of the somatosensory cortex, which were fully impregnated and free of neighboring cells or cellular debris, were randomly selected for analysis (Supplementary Fig. 16 ). Golgi-stained coronal sections containing medial amygdala and somatosensory cortical area were visualized under Olympus BX51 microscope. Neuronal tracing was carried out with the help of Neurolucida and Neuroexplorer software (MicroBright Field Inc., Williston, USA). Up to three pyramidal cells in the MeA and four pyramidal cells in the somatosensory cortex per section over 3 sections (9 (MeA) and 12 (cortex) cells per animal respectively) were sampled 88 , 89 . For Sholl analysis 90 , concentric circles were placed at 10 µm intervals starting from the center of the cell body and the parameters i) total dendritic length (sums of the length of individual branches) of apical and basal dendrites of pyramidal cells and ii) number of spines (protrusions in direct contact with the primary dendrite) and their density (number of spines per 10 µm) were recorded.
Neuron selection criteria: Neurons were selected based on the following criteria. They had to be fully stained, and the cell body had to be in the middle third of the section thickness. The dendrites of the neuron had to be unobscured by the other nearby neuron. Also, neurons had to possess tapering of the majority of the dendrites towards their ends. Representative images of neurons from vehicle and BPA-exposed adult mice can be found in Supplementary Fig. 17 .
Visualizing c-Fos activation to conspecific exposure (amygdala)
Stranger exposure paradigm procedure.
Cyp19-EGFP mice of both sexes as well as male ArKO mice, together with male WT littermates were utilized in this study. Mice had not undergone any other behavior testing. All test mice were acclimatized to the testing room in individual cages for three nights prior to testing. All mice were age P24 on the day of testing, which was performed between 10 am-2 pm. Testing was performed in a dedicated room for mouse behavior studies and no other animals were present in the room at the time of acclimatization and testing. The temperature of the room was maintained at approximately 21 °C.
On the day of testing, each mouse cage containing the isolated test mouse was placed on a stage (a trolley). The lid containing food and water was removed and immediately following, a sex-/age-matched C57Bl/6 J stranger mouse or a novel object (new 1 mL syringe) was placed into the cage and a clean, empty lid was placed on the top. New gloves were used to handle each syringe to avoid transferring another mouse’s olfactory signature to it. The 10 min trial began as soon as the cage lid was shut. After 10 min had elapsed, the stranger or the novel object was removed, the test mouse with home cage was returned to its original location with the original cage lid with food and water for 2 hours prior to perfusion. Once it was established that there was a difference in c-fos expression between stranger exposure and novel object exposure in the medial amygdala, BPA and vehicle exposed (ED10.5-14.5) Cyp19-EGFP mice as well as estrogen and sham pallet treated ArKO and WT mice were exposed to an age and sex matched stranger as described above. C-fos expression was quantified in male mice only.
Histology and stereological analysis methods can be found in the Supplementary Methods.
Neuron count brain collection, staining, brain region delineation and stereology can be found in the Supplementary Methods.
Electrophysiological studies
Microelectrode array electrophysiology.
Male mice aged 8 weeks weighing between 15 and 20 g were used for this study. They had not undergone any behavioral testing prior to electrophysiology. We studied synaptic activity parameters such as the Input/Output (I/O) curve. Stimulation of the glutamatergic synapses terminate in the basolateral amygdala (BLA) and the basomedial amygdala (BMA), which were integrated with multiple inputs that compute to produce an output (field excitatory postsynaptic potential, fEPSP). I/O curve serves as an index of synaptic excitability of large neuronal populations. Mice were anesthetized with isoflurane (IsoFlo TM , Abbott Laboratories, Victoria, Australia) and decapitated. The whole brains were quickly removed and placed in ice-cold, oxygenated (95% O 2 , 5% CO 2 ) cutting solution (composition in mmol/L: 206 sucrose, 3 KCl, 0.5 CaCl 2 , 6 MgCl 2 -H 2 O, 1.25 NaH 2 PO 4 , 25 NaHCO 3, and 10.6 D-glucose). Coronal brain amygdala slices (300 µm) were prepared with a VT 1200 S tissue slicer (Leica) and quickly transferred to 34 o C carbogen bubbled artificial CSF (aCSF) (composition in mmol/L: 126 NaCl, 2.5 KCl, 2.4 CaCl 2 , 1.36 MgCl 2 -H 2 O, 1.25 NaH 2 PO 4 , 25 NaHCO 3, and 10 D-glucose) for 30 min. After further recovery of 1 h equilibrium in oxygenated aCSF at room temperature, the slices were transferred to a submission recording chamber, an MEA chip with 60 electrodes spaced 200 μm apart (60 MEA 200/30 iR-Ti: MCS GnbH, Reutlingen, Germany). The slice was immobilized with a harp grid (ALA Scientific Instruments, New York, USA) and was continuously perfused with carbogenated aCSF (3 mL/min at 32 °C). fEPSPs produced in BLA and BMA were by stimulation of a randomly chosen electrode surrounding the target area with a biphasic voltage waveform (100 μs) at intermediated voltage intensity. The electrode could only be chosen if it produced a fair number of fEPSPs in the surrounding recording electrodes. The width of the EPSP wave ranged from 20 to 30 ms was selected. We chose slices where BLA and BMA were greatly represented according to Allen Mouse Brain Atlas 91 . Care was taken to choose the stimulating electrode in the same region from one slice to the other. The peak-to-peak amplitude of fEPSP in BLA and BMA was recorded by a program of LTP-Director and analyzed using LTP-Analyzer (MCS GnbH, Reutlingen, Germany).
Electrocorticogram (ECoG)
Electrocorticogram recordings.
Male mice aged 8 weeks were used for this study. Mice had not undergone any behavioral testing prior to ECoG recording. For ECoG, surgeries were performed as previously described 92 . Mice were anesthetized with 1–3% isoflurane and two epidural silver ‘ball’ electrodes implanted on each hemisphere of the skull. Electrodes were placed 3 mm lateral of the midline and 0.5 mm, caudal from bregma. A ground electrode was placed 2.5 mm rostral from bregma and 0.5 mm lateral from the midline. Mice were allowed to recover for at least 48 hours after surgery. ECoGs were continuously recorded in freely moving mice for a 4–6-hour period during daylight hours following a standard 30-min habituation period. Signals were band-pass filtered at 0.1 to 40 Hz and sampled at 1 kHz using the Pinnacle EEG/EMG tethered recording system (Pinnacle Technology Inc, KS). Power spectrums were calculated using Hann window with a resolution of 1 Hz using Sirenia Pro analysis software (Pinnacle Technology Inc) on stable 30-min periods of ECoG recordings.
Primary Cortical Cultures
Neuroprotective effect of 10hda against injury induced by bpa on embryonic mouse cortical neurons.
Primary cortical neurons were obtained from male Cyp19 -EGFP mouse embryos at gestational day 15.5. Embryos were genotyped for SRY to determine sex, and only male embryos were used. Cells were seeded in 24-well plates containing 12 mm glass coverslips, coated with 100 µg/mL poly D-lysine to a density of ~0.45 x 10 6 cells/well and incubated in a humidified CO 2 incubator (5% CO 2 , 37 °C). Cells were pre-treated with vehicle (DMSO), 1 mM 10HDA (Matreya, PA,USA), 25 nM BPA and 1 mM 10HDA with 25 nM BPA. For each group, 10 neurons were measured, and the experiments were duplicated. Each replicate was from a separate culture.
Cells were fixed in 4% paraformaldehyde and stained with mouse anti-βIII tubulin monoclonal primary antibody (1:1000; cat #ab41489, Abcam, United Kingdom) and goat anti-mouse secondary antibody, Alexa Fluor 488, (1:2000; cat#A11017; Invitrogen, USA) to label neuronal cells. Aromatase was stained using Rabbit anti-aromatse Antibody (1:2000 cat# A7981; Sigma Aldrich, St. Louis, MO, USA) and donkey anti-rabbit Alexa594 (1:2000; cat# A-21207; Invitrogen, USA). Cell nucleus was stained with Hoechst 33258 solution (Sigma 94403 (2 µg/mL)). Images were captured using an Olympus IX51 microscope (X40 objective). Neurites were quantified using Neurolucida and Neuroexplorer software (MicroBright Field Inc, Williston, USA) as described in the neuron tracing section.
RNA extraction
Total RNA was extracted using PARIS kit (cat#: AM1921, Invitrogen™PARIS™ Kit) according to the protocol supplied by the manufacturer. cDNA libraries were generated using the SureSelect.
Strand-Specific RNA Library Prep for Illumina Multiplexed Sequencing kit (Agilent Technologies, CA, USA), according to manufacturer’s instructions.
In vivo effects of BPA on Fetal brain cortical RNA seq
Pregnant Cyp19-EGFP dams were injected subcutaneously with BPA or vehicle ED10.5-14.5 as described in previous section, and culled on ED15.5 by isoflurane overdosed. Fetuses were harvested and placed in chilled PBS. Embryo brain cortical tissue was dissected from fetuses, snap frozen in liquid nitrogen and stored in −80°C until RNA extraction. The sex of fetuses was determined by visual assessment of the gonads and Sry (a male-specific gene) genotyping. Each RNA seq run, 6 cDNA libraries (derived from total RNA samples with 3 biological samples per group), were analyzed by MidSeq Nano run, 50 bp, Single end read on the Illumina platform. Because of undetectable levels of Cyp19a1 RNA in the fetal brain, Cyp19a1 RNA levels were not included. This is consistent in that Aromatase+ cells represent <0.05% of neurons in the adult mouse brain 93 . Subsequent in vivo transcriptomic analyses were completed in males only. Read quality was then assessed with FastQC. The sequence reads were then aligned against the Mus musculus genome (Build version GRCm38). The Tophat aligner (v2.0.14) was used to map reads to the genomic sequences. Sequencing data were then summarized into reads per transcript using Feature counts 94 . The transcripts were assembled with the StringTie tool v1.2.4 using the reads alignment with Mus_musculus.GRCm38 and reference annotation based assembly option (RABT) using the Gencode gene models for the mouse GRCm38/mm10 genome build. Normalisation and statistical analysis on the count data were executed using EdgeR (version edgeR_3.14.2 in R studio, R version 3.14.2). The data were scaled using trimmed mean of M-values (TMM) 95 and differentially expressed genes between all treatment group (Benjamini–Hochberg false discovery rate >0.1). Differentially expressed genes (DEGs) were identified by comparing mice exposed to 50 μg/kg/day BPA with those exposed to the vehicle.
In vitro effects of 10HDA in primary cell culture RNASeq
Primary brain cortical neurons were obtained from C57BL/6 mouse embryos at GD 15.5. Neuronal cell cultures were treated with vehicle (DMSO) or 1 mM 10HDA (Matreya, PA,USA) as described above. The libraries were sequenced with 50 bp single end reads using an Illumina Hiseq and read quality assessed using FastQC. Untrimmed reads were aligned to mouse mm10 genome using Subjunc aligner (version 1.4.4) within the Subread package 96 . Sequencing data were then summarized into reads per transcript using Feature counts 94 and the Gencode gene models for the mouse GRCm38/mm10 genome build (August 2014 freeze) 97 . Normalisation and statistical analysis on the count data were executed using EdgeR (version edgeR_3.4.2 in R studio, R version 3.0.2) 98 after removing features with less than 10 counts per million for at least 3 of the samples. The data were scaled using trimmed mean of M-values (TMM) 95 and differentially expressed genes between all treatment group (Benjamini–Hochberg false discovery rate >0.1). Annotation was added using the ensemble mouse gene annotation added using bioMart package 99 . Differentially expressed genes were identified by comparing cells exposed to 10HDA with those exposed to the vehicle.
Pathway analysis
The BPA and 10HDA differential expression data for enriched pathways were analyzed using Ingenuity (QIAGEN) and tested against the Canonical Pathway Library, Brain Diseases and Functions Library and the Brain Disorders pathway libraries. We included the top 8 Canonical pathways. Then we included only pathways which were p < 0.05 in both the BPA and the 10HDA data for the Brain Diseases and Functions pathway libraries, and included all P -values for the Brain Disorders pathway library ( p > 0.05 are in gray).
An additional analysis of the gene expression data was performed using the c lusterProfileR R package 100 , which provides a range of statistical tests to detect pathways from a query gene set. The test used here was Gene Set Enrichment Analysis (GSEA) 101 , and the genes were tested against the Gene Ontology pathway database (specifically, GO: Biological Process) 102 .
Computational Molecular Docking
The DockThor molecular docking platform 103 was used to assess binding affinities between estrogen receptor beta (encoded by ESR2 gene) and the ligands 17-beta estradiol (E2; the native ligand), BPA, and 10HDA.DockThor takes as input 3D molecular structures for a putative receptor-ligand pair and employs a genetic-algorithm-based optimization strategy to identify optimal binding position within a specified search region. The crystal structures of estrogen receptor alpha (Erα) and beta (Erβ) were sourced from the Protein Data Bank (PDB) with respective PDB IDs: Erα - 5KRI and Erβ - 1YYE. For each ligand, the search grid was restricted to the known estrogen receptor beta ligand-binding domain, centered at x = 30, y = 35, z = 40, with total grid size of x = 25, y = 28 and z = 22. Default settings were used for the optimization procedure.
Statistical analysis
Researchers were blind to treatment during the conduct of the experiment and the outcome assessment but not during statistical analysis.
Mean, standard deviation (SD), and standard error of the mean (SEM) were calculated with GraphPad Prism version 9.4 (GraphPad Software).
Data were tested for equal variances and normality using the Shapiro Wilk test. As electrophysiology, Golgi staining and primary cell culture experiments utilized several data points per animal, observations were not independent, and this non-independence was accounted for in our analyses. We used generalized estimating equations (GEEs) in R version 4.1.2. in a marginal modeling approach that estimates population-averaged effects while treating the covariance structure as a nuisance. We specified the covariance structure as exchangeable (that is, assumed equal correlation between pairs of measurements on the same animal). Given the small number of clusters (i.e. animals), bootstrapped standard errors were estimated using 200 repeats to maintain a conservative type 1 error rate 104 . An interaction term was added to the amygdala Golgi staining study to assess a sex * genotype or sex * BPA exposure interaction.
Where data were normally distributed, parametric tests were conducted. For more than two groups, a one-way ANOVA was conducted with Holm-Sidak post hoc FDR correction, with alpha set to 0.05. Otherwise, unpaired two-tailed Student’s t -tests were used to compare two variables. In cases where normality was not assumed, a Mann-Whitney (comparing two groups) or Kruskal-Wallis with Dunn’s post hoc (comparing three or more groups) was used. In the case of the three-chamber data, a two-way mixed ANOVA was used to assess group x cage side interaction (stranger cage interaction zone vs empty cage interaction zone) with post hoc testing adjusted by the Holm-Sidak method.
Comparisons made are indicated on the Figure legends, and p -values < 0.05 were considered significant. All tests were two-sided (two-tailed) where applicable.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The BIS data including all data used in this paper are available under restricted access for participant privacy. Access can be obtained by request through the BIS Steering Committee by contacting Anne-Louise Ponsonby, The Florey institute of Neuroscience and Mental Health, [email protected]. Requests to access cohort data will be responded to within two weeks. Requests are then considered on scientific and ethical grounds and, if approved, provided under collaborative research agreements. Deidentified cohort data can be provided in Stata or CSV format. Additional project information, including cohort data description and access procedure, is available at the project’s website https://www.barwoninfantstudy.org.au . Source data underlying Figs. 1 – 6 , 8 – 10 and Supplementary Figs. 2 , 3 – 7 , 14 have been provided as a Source Data file with this paper. The RNAseq data discussed in this publication have been deposited in NCBI’s Gene Expression Omnibus 105 and are accessible through GEO Series accession numbers; fetal brain expression with and without prenatal BPA exposure, GSE266401 and primary cortical culture treated with and without 10HDA, GSE266400 Source data are provided with this paper.
American Psychiatric Association aAPAD-TF. Diagnostic and Statistical Manual of Mental Disorders: DSM-5 (American Psychiatric Association, 2013).
Maenner, M. et al. Prevalence of autism spectrum disorder among children aged 8 years—autism and developmental disabilities monitoring network, 11 sites, United States, 2016. MMWR Surveill. Summ. 69 , 1–12 (2020).
Article PubMed PubMed Central Google Scholar
Li, Q. et al. Prevalence of autism spectrum disorder among children and adolescents in the United States From 2019 to 2020. JAMA Pediatrics 176 , 943–945 (2022).
Atladottir, H. O. et al. The increasing prevalence of reported diagnoses of childhood psychiatric disorders: a descriptive multinational comparison. Eur. Child Adolesc. Psychiatry 24 , 173–183 (2015).
Article PubMed Google Scholar
Lai, M.-C. & Lombardo, M. V. Baron-Cohen S. autism. Lancet 383 , 896–910 (2014).
Boon, W. C., Chow, J. D. & Simpson, E. R. The multiple roles of estrogens and the enzyme aromatase. Prog. Brain Res. 181 , 209–232 (2010).
Article CAS PubMed Google Scholar
Harada, N. & Honda, S.-i Analysis of spatiotemporal regulation of aromatase in the brain using transgenic mice. J. Steroid Biochem Mol. Biol. 95 , 49–55 (2005).
Tan, W., Zhu, Z., Ye, L. & Leung, L. K. Methylation dictates PI.f-specific CYP19 transcription in human glial cells. Mol. Cell Endocrinol. 452 , 131–137 (2017).
Roselli, C. E., Liu, M. & Hurn, P. D. Brain aromatization: classic roles and new perspectives. Semin Reprod. Med. 27 , 207–217 (2009).
Article CAS PubMed PubMed Central Google Scholar
Ruiz-Palmero, I. et al. Oestradiol synthesized by female neurons generates sex differences in neuritogenesis. Sci. Rep. 6 , 31891 (2016).
Article ADS CAS PubMed PubMed Central Google Scholar
Stanić, D. et al. Characterization of aromatase expression in the adult male and female mouse brain. I. Coexistence with oestrogen receptors α and β, and androgen receptors. PLoS One 9 , e90451 (2014).
Article ADS PubMed PubMed Central Google Scholar
Takahashi, K. et al. Association between aromatase in human brains and personality traits. Sci. Rep. 8 , 16841 (2018).
Baron-Cohen, S. The extreme male brain theory of autism. Trends Cogn. Sci. 6 , 248–254 (2002).
Crider, A., Thakkar, R., Ahmed, A. O. & Pillai, A. Dysregulation of estrogen receptor beta (ERβ), aromatase (CYP19A1), and ER co-activators in the middle frontal gyrus of autism spectrum disorder subjects. Mol. Autism 5 , 46–46 (2014).
Sarachana, T. & Hu, V. W. Genome-wide identification of transcriptional targets of RORA reveals direct regulation of multiple genes associated with autism spectrum disorder. Mol. Autism 4 , 14 (2013).
Santangeli, S. et al. Effects of BPA on female reproductive function: the involvement of epigenetic mechanism. Gen. Comp. Endocrinol. 245 , 122–126 (2017).
Ali, A. A. et al. Developmental vitamin D deficiency increases foetal exposure to testosterone. Mol. Autism 11 , 96 (2020).
Andrade, A. J., Grande, S. W., Talsness, C. E., Grote, K. & Chahoud, I. A dose-response study following in utero and lactational exposure to di-(2-ethylhexyl)-phthalate (DEHP): non-monotonic dose-response and low dose effects on rat brain aromatase activity. Toxicology 227 , 185–192 (2006).
Moosa, A., Shu, H., Sarachana, T. & Hu, V. W. Are endocrine disrupting compounds environmental risk factors for autism spectrum disorder? Hormones Behav. 101 , 13–21 (2018).
Article CAS Google Scholar
Ejaredar, M., Lee, Y., Roberts, D. J., Sauve, R. & Dewey, D. Bisphenol A exposure and children’s behavior: a systematic review. J. Expo. Sci. Environ. Epidemiol. 27 , 175–183 (2017).
Chen, D. et al. Bisphenol analogues other than BPA: environmental occurrence, human exposure, and toxicity—a review. Environ. Sci. Technol. 50 , 5438–5453 (2016).
Article ADS CAS PubMed Google Scholar
Hansen, J. B. et al. Prenatal exposure to bisphenol A and autistic- and ADHD-related symptoms in children aged 2 and 5 years from the Odense Child Cohort. Environ. Health 20 , 24 (2021).
Lim, Y. H. et al. Prenatal and postnatal bisphenol A exposure and social impairment in 4-year-old children. Environ. Health 16 , 79 (2017).
Ferri, S. L., Abel, T. & Brodkin, E. S. Sex differences in autism spectrum disorder: a review. Curr. Psychiatry Rep. 20 , 9 (2018).
Suzuki, K. M. et al. Estrogenic activities of fatty acids and a sterol isolated from royal jelly. Evid. Based Complement. Alternat. Med. 5 , 295–302 (2008).
Pirgon, O., Atar, M., Çiriş, M. & Sever, M. Effects of royal jelly supplementation on growth plate zones and longitudinal growth in young rats. Mellifera 19 , 1–13 (2019).
Google Scholar
Hattori, N., Nomoto, H., Fukumitsu, H., Mishima, S. & Furukawa, S. Royal jelly and its unique fatty acid, 10-hydroxy-trans-2-decenoic acid, promote neurogenesis by neural stem/progenitor cells in vitro. Biomed. Res. 28 , 261–266 (2007).
Vuillermin, P. et al. Cohort profile:the Barwon Infant Study. Int J. Epidemiol. 44 , 1148–1160 (2015).
Achenbach T. M. & Rescorla L. A. Manual for the ASEBA Preschool Forms & Profiles (University of Vermont, Research Center for Children, Youth, & Families, 2000).
Pham, C. et al. Early life environmental factors associated with autism spectrum disorder symptoms in children at age 2 years: a birth cohort study. Autism.: Int. J. Res. Pract. 26 , 1864–1881 (2022).
Article Google Scholar
Kidokoro, K. et al. Association between CYP19A1 polymorphisms and sex hormones in postmenopausal Japanese women. J. Hum. Genet. 54 , 78–85 (2009).
Braun, P. R. et al. Genome-wide DNA methylation comparison between live human brain and peripheral tissues within individuals. Transl. Psychiatry 9 , 47 (2019).
Kundakovic, M. et al. DNA methylation of BDNF as a biomarker of early-life adversity. Proc. Natl Acad. Sci. USA 112 , 6807–6813 (2015).
Panickar, K. S., Guan, G., King, M. A., Rajakumar, G. & Simpkins, J. W. 17beta-estradiol attenuates CREB decline in the rat hippocampus following seizure. J. Neurobiol. 33 , 961–967 (1997).
Solum, D. T. & Handa, R. J. Estrogen regulates the development of brain-derived neurotrophic factor mRNA and protein in the rat hippocampus. J. Neurosci. 22 , 2650–2659 (2002).
EPA USEPAU. Bisphenol A; CASRN 80-05-7. Integrated Risk Information System (IRIS) Chemical Assessment Summary (National Center for Environmental Assessment, 1988).
U.S. Food & Drug Administration. Bisphenol A (BPA): Use in Food Contact Application (U.S. Food & Drug Administration, 2023). [cited 2024 Jul]. Available from: https://www.fda.gov/food/food-additives-petitions/bisphenol-bpa-use-food-contact-application .
Authority, E. F. S. Opinion of the Scientific Panel on food additives, flavourings, processing aids and materials in contact with food (AFC) related to 2, 2‐BIS (4‐HYDROXYPHENYL) PROPANE. EFSA J. 5 , 428 (2007).
Semple, B. D., Dixit, S., Shultz, S. R., Boon, W. C. & O’Brien, T. J. Sex-dependent changes in neuronal morphology and psychosocial behaviors after pediatric brain injury. Behav. Brain Res. 319 , 48–62 (2017).
Soma, M. et al. Development of the mouse amygdala as revealed by enhanced green fluorescent protein gene transfer by means of in utero electroporation. J. Comp. Neurol. 513 , 113–128 (2009).
Fisher, C. R., Graves, K. H., Parlow, A. F. & Simpson, E. R. Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc. Natl Acad. Sci. USA 95 , 6965–6970 (1998).
Hill, R. A. et al. Estrogen deficient male mice develop compulsive behavior. Biol. Psychiatry 61 , 359–366 (2007).
Avino, T. A. et al. Neuron numbers increase in the human amygdala from birth to adulthood, but not in autism. Proc. Natl Acad. Sci. USA 115 , 3710–3715 (2018).
Cai, S., Wang, X., Yang, F., Chen, D. & Huang, L. Differences in brain structural covariance network characteristics in children and adults with autism spectrum disorder. Autism Res. 14 , 265–275 (2021).
Keshavarzi, S., Sullivan, R. K. P., Ianno, D. J. & Sah, P. Functional properties and projections of neurons in the medial amygdala. J. Neurosci. 34 , 8699–8715 (2014).
Alia-Klein, N. et al. Human cognitive ability is modulated by aromatase availability in the brain in a sex-specific manner. Front Neurosci. 14 , 565668 (2020).
Biegon, A. et al. Relationship of estrogen synthesis capacity in the brain with obesity and self-control in men and women. Proc. Natl Acad. Sci. USA 117 , 22962–22966 (2020).
Sato, W. & Uono, S. The atypical social brain network in autism: advances in structural and functional MRI studies. Curr. Opin. Neurol. 32 , 617–621 (2019).
Herrington, J. D., Miller, J. S., Pandey, J. & Schultz, R. T. Anxiety and social deficits have distinct relationships with amygdala function in autism spectrum disorder. Soc. Cogn. Affect Neurosci. 11 , 907–914 (2016).
Krukoff, T. L. C-fos expression as a marker of functional activity in the brain. in Cell Neurobiology Techniques. Neuromethods . Vol. 33 213–230 (Humana Press, 1999).
Stoner, R. et al. Patches of disorganization in the neocortex of children with autism. N. Engl. J. Med. 370 , 1209–1219 (2014).
Anthoni, H. et al. The aromatase gene CYP19A1: several genetic and functional lines of evidence supporting a role in reading, speech and language. Behav. Genet. 42 , 509–527 (2012).
Wang, J. et al. Resting state EEG abnormalities in autism spectrum disorders. J. Neurodev. Disord. 5 , 24 (2013).
Matuszczak, E., Komarowska, M. D., Debek, W. & Hermanowicz, A. The impact of Bisphenol A on fertility, reproductive system, and development: a review of the literature. Int. J. Endocrinol. 2019 , 4068717 (2019).
Moutsatsou, P. et al. Fatty acids derived from royal jelly are modulators of estrogen receptor functions. PLoS ONE 5 , e15594 (2010).
Rubin B. S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid. Biochem. Mol. Biol . 127 , 27–34 (2011).
DeRosa, B. A. et al. Convergent pathways in idiopathic autism revealed by time course transcriptomic analysis of patient-derived neurons. Sci. Rep. 8 , 8423 (2018).
Andrews, S. V. et al. Case-control meta-analysis of blood DNA methylation and autism spectrum disorder. Mol. Autism 9 , 40 (2018).
Thongkorn, S. et al. Sex differences in the effects of prenatal bisphenol A exposure on autism-related genes and their relationships with the hippocampus functions. Sci. Rep. 11 , 1241 (2021).
VanderWeele, T. J. Explanation in causal inference: developments in mediation and interaction. Int J. Epidemiol. 45 , 1904–1908 (2016).
PubMed PubMed Central Google Scholar
Gerona, R., vom Saal, F. S. & Hunt, P. A. BPA: have flawed analytical techniques compromised risk assessments? Lancet Diabetes Endocrinol. 8 , 11–13 (2020).
Bolognesi, C. et al. Scientific Opinion on the risks to public health related to the presence of bisphenol A (BPA) in foodstuffs: Executive summary. efsa J. (2015).
Authority E. F. S. Re-evaluation of the Risks to Public Health Related to the Presence of Bisphenol A (BPA) in foodstuffs (European Food Safety Authority, 2021).
EFSA Panel on Food Contact Materials E, et al. Re-evaluation of the risks to public health related to the presence of bisphenol A (BPA) in foodstuffs. EFSA J. 21 , e06857 (2023).
Gao, H. et al. Cumulative risk assessment of phthalates associated with birth outcomes in pregnant Chinese women: a prospective cohort study. Environ. Pollut. 222 , 549–556 (2017).
Welch, C., Mulligan, K. & Does Bisphenol, A. Confer risk of neurodevelopmental disorders? what we have learned from developmental neurotoxicity studies in animal models. Int J. Mol. Sci. 23 , 2894 (2022).
Stein, T. P., Schluter, M. D., Steer, R. A., Guo, L. & Ming, X. Bisphenol A exposure in children with autism spectrum disorders. Autism Res. 8 , 272–283 (2015).
Rudel, R. A. et al. Food Packaging and Bisphenol A and Bis(2-Ethyhexyl) Phthalate Exposure: Findings from a Dietary Intervention. Environ. Health Perspect. 119 , 914–920 (2011).
Geueke, B. et al. Systematic evidence on migrating and extractable food contact chemicals: Most chemicals detected in food contact materials are not listed for use. Criti. Rev. Food Sci. Nutrition 63, 9425–9435 (2022).
Arbuckle, T. E. et al. Exposure to free and conjugated forms of bisphenol a and triclosan among pregnant women in the MIREC cohort. Environ. Health Perspect. 123 , 277–284 (2015).
Caporale, N. et al. From cohorts to molecules: adverse impacts of endocrine disrupting mixtures. Science 375 , eabe8244 (2022).
Hameed, R. A., Ahmed, E. K., Mahmoud, A. A. & Atef, A. A. G protein-coupled estrogen receptor (GPER) selective agonist G1 attenuates the neurobehavioral, molecular and biochemical alterations induced in a valproic acid rat model of autism. Life Sci. 328 , 121860 (2023).
Heffernan, A. L. et al. Harmonizing analytical chemistry and clinical epidemiology for human biomonitoring studies. A case-study of plastic product chemicals in urine. Chemosphere 238 , 124631 (2020).
Burugupalli, S. et al. Ontogeny of circulating lipid metabolism in pregnancy and early childhood—a longitudinal population study. Elife 11 , e72779 (2022).
McCarthy, S. et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 48 , 1279–1283 (2016).
Ponsonby, A. L. et al. Prenatal phthalate exposure, oxidative stress-related genetic vulnerability and early life neurodevelopment: a birth cohort study. Neurotoxicology 80 , 20–28 (2020).
Wang, Y. et al. A methodological pipeline to generate an epigenetic marker of prenatal exposure to air pollution indicators. Epigenetics 17 , 32–40 (2022).
Perera, F. et al. Prenatal bisphenol a exposure and child behavior in an inner-city cohort. Environ. Health Perspect. 120 , 1190–1194 (2012).
Hunt, P. A., Vom Saal, F. S., Stahlhut, R. & Gerona, R. BPA and risk assessment—Authors’ reply. Lancet Diabetes Endocrinol. 8 , 271–272 (2020).
Burke James, F., Sussman Jeremy, B., Kent David, M. & Hayward Rodney, A. Three simple rules to ensure reasonably credible subgroup analyses. BMJ 351 , h5651 (2015).
Ponsonby, A.-L. Reflection on modern methods: building causal evidence within high-dimensional molecular epidemiological studies of moderate size. Int J. Epidemiol. 50 , 1016–1029 (2021).
Team RC. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2013).
StataCorp. Stata Statistical Software: Release 14 (StataCorp LP, 2015).
Thongkorn, S. et al. Investigation of autism-related transcription factors underlying sex differences in the effects of bisphenol A on transcriptome profiles and synaptogenesis in the offspring hippocampus. Biol. Sex. Differ. 14 , 8 (2023).
Nadler, J. J. et al. Automated apparatus for quantitation of social approach behaviors in mice. Genes, brain, Behav. 3 , 303–314 (2004).
Moy, S. S. et al. Sociability and preference for social novelty in five inbred strains: an approach to assess autistic-like behavior in mice. Genes, Brain, Behav. 3 , 287–302 (2004).
Champagne, D. L. et al. Maternal care and hippocampal plasticity: evidence for experience-dependent structural plasticity, altered synaptic functioning, and differential responsiveness to glucocorticoids and stress. J. Neurosci. 28 , 6037–6045 (2008).
Wang, I. T., Reyes, A. R. & Zhou, Z. Neuronal morphology in MeCP2 mouse models is intrinsically variable and depends on age, cell type, and Mecp2 mutation. Neurobiol. Dis. 58 , 3–12 (2013).
Kelly, E. A., Opanashuk, L. A. & Majewska, A. K. The effects of postnatal exposure to low-dose bisphenol-A on activity-dependent plasticity in the mouse sensory cortex. Front. Neuroanat. 8 , 117 (2014).
Sholl, D. A. Dendritic organization in the neurons of the visual and motor cortices of the cat. J. Anat. 87 , 387–406 (1953).
ADS CAS PubMed PubMed Central Google Scholar
Allen Institute for Brain Science AMBA. https://mouse.brain-map.org/static/atlas (2004).
Tan, H. O. et al. Reduced cortical inhibition in a mouse model of familial childhood absence epilepsy. Proc. Natl Acad. Sci. USA 104 , 17536–17541 (2007).
Unger, E. K. et al. Medial amygdalar aromatase neurons regulate aggression in both sexes. Cell Rep. 10 , 453–462 (2015).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30 , 923–930 (2013).
Robinson, M. D. & Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 11 , R25 (2010).
Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41 , e108 (2013).
Harrow, J. et al. GENCODE: producing a reference annotation for ENCODE. Genome Biol. 7 , S4.1–9 (2006).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26 , 139–140 (2010).
Durinck, S., Spellman, P. T., Birney, E. & Huber, W. Mapping identifiers for the integration of genomic datasets with the R/Bioconductor package biomaRt. Nat. Protoc. 4 , 1184–1191 (2009).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation 2 , 100141 (2021).
CAS PubMed PubMed Central Google Scholar
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102 , 15545–15550 (2005).
The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 47 , D330–d338 (2019).
Santos, K. B., Guedes, I. A., Karl, A. L. M. & Dardenne, L. E. Highly flexible ligand docking: benchmarking of the DockThor Program on the LEADS-PEP protein–peptide data set. J. Chem. Inf. Model 60 , 667–683 (2020).
Leyrat, C., Morgan, K. E., Leurent, B. & Kahan, B. C. Cluster randomized trials with a small number of clusters: which analyses should be used? Int J. Epidemiol. 47 , 321–331 (2018).
Edgar, R., Domrachev, M. & Lash, A. E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 30 , 207–210 (2002).
Xiao, X. et al. Bisphenol AP is anti-estrogenic and may cause adverse effects at low doses relevant to human exposure. Environ. Pollut. 242 , 1625–1632 (2018).
Download references
Acknowledgements
The authors thank the BIS participants for their generous contribution to this project. The authors also thank current and past cohort staff. The establishment work and infrastructure for the BIS was provided by the Murdoch Children’s Research Institute, Deakin University, and Barwon Health, supported by the Victorian Government’s Operational Infrastructure Program. We thank all the children and families participating in the study, and the BIS fieldwork team. We acknowledge Barwon Health, Murdoch Children’s Research Institute, and Deakin University for their support in the development of this research. We thank Dr Shanie Landen for statistical advice, and Alex Eisner for independent statistical review of the analyses in the manuscript. We thank Soumini Vijayay and Kristie Thompson for human BPA lab measurement and Dr Steve Cheung for assistance preparing the primary cortical culture. We thank Chitra Chandran, Georgia Cotter, Stephanie Glynn, Oliver Wood and Janxian Ng for manuscript preparation. Manuscript editor Julian Heng (Remotely Consulting, Australia) provided professional editing of this article. This multimodal project was supported by funding from the Minderoo Foundation. Funding was also provided by the National Health and Medical Research Council of Australia (NHMRC), the NHMRC-EU partnership grant for the ENDpoiNT consortium, the Australian Research Council, the Jack Brockhoff Foundation, the Shane O’Brien Memorial Asthma Foundation, the Our Women’s Our Children’s Fund Raising Committee Barwon Health, The Shepherd Foundation, the Rotary Club of Geelong, the Ilhan Food Allergy Foundation, GMHBA Limited, Vanguard Investments Australia Ltd, and the Percy Baxter Charitable Trust, Perpetual Trustees, Fred P Archer Fellowship; the Scobie Trust; Philip Bushell Foundation; Pierce Armstrong Foundation; The Canadian Institutes of Health Research; BioAutism, William and Vera Ellen Houston Memorial Trust Fund, Homer Hack Research Small Grants Scheme and the Medical Research Commercialisation Fund. This work was also supported by Ms. Loh Kia Hui. This project received funding from a NHMRC-EU partner grant with the European Union’s Horizon 2020 Research and Innovation Programme, under Grant Agreement number: 825759 (ENDpoiNTs project). This work was also supported by NHMRC Investigator Fellowships (GTN1175744 to D.B., APP1197234 to A.-L.P., and GRT1193840 to P.S.). The study sponsors were not involved in the collection, analysis, and interpretation of data; writing of the report; or the decision to submit the report for publication.
Author information
Nhi Thao Tran
Present address: The Ritchie Centre, Department of Obstetrics and Gynaecology, School of Clinical Sciences, Monash University, Clayton, Australia
These authors contributed equally: Christos Symeonides, Kristina Vacy.
These authors jointly supervised this work: Anne-Louise Ponsonby, Wah Chin Boon.
Authors and Affiliations
Minderoo Foundation, Perth, Australia
- Christos Symeonides
Murdoch Children’s Research Institute, Parkville, Australia
Christos Symeonides, Toby Mansell, Martin O’Hely, Boris Novakovic, David Burgner, Mimi L. K. Tang, Richard Saffery, Peter Vuillermin, Fiona Collier, Anne-Louise Ponsonby, Sarath Ranganathan, Lawrence Gray & Anne-Louise Ponsonby
Centre for Community Child Health, Royal Children’s Hospital, Parkville, Australia
Christos Symeonides, Sarath Ranganathan & Anne-Louise Ponsonby
The Florey Institute of Neuroscience and Mental Health, Parkville, Australia
Kristina Vacy, Sarah Thomson, Sam Tanner, Hui Kheng Chua, Shilpi Dixit, Jessalynn Chia, Nhi Thao Tran, Sang Eun Hwang, Feng Chen, Tae Hwan Kim, Christopher A. Reid, Anthony El-Bitar, Gabriel B. Bernasochi, Anne-Louise Ponsonby, Anne-Louise Ponsonby & Wah Chin Boon
School of Population and Global Health, The University of Melbourne, Parkville, Australia
Kristina Vacy
The Hudson Institute of Medical Research, Clayton, Australia
Hui Kheng Chua & Yann W. Yap
Department of Pediatrics, The University of Melbourne, Parkville, Australia
Toby Mansell & David Burgner
School of Medicine, Deakin University, Geelong, Australia
Martin O’Hely, Boris Novakovic, Chloe J. Love, Peter D. Sly, Peter Vuillermin, Fiona Collier & Lawrence Gray
Columbia Center for Children’s Environmental Health, Columbia University, New York, NY, USA
Julie B. Herbstman, Shuang Wang & Jia Guo
Department of Environmental Health Sciences, Columbia University, New York, NY, USA
Julie B. Herbstman
Department of Biostatistics, Columbia University, New York, NY, USA
Shuang Wang & Jia Guo
Department of Anatomy and Developmental Biology, Monash University, Clayton, Australia
Kara Britt & Vincent R. Harley
Breast Cancer Risk and Prevention Laboratory, Peter MacCallum Cancer Centre, Melbourne, Australia
Sir Peter MacCallum Department of Oncology, The University of Melbourne, Melbourne, Australia
Faculty Medicine, Dentistry & Health Sciences, University of Melbourne, Parkville, Australia
Gabriel B. Bernasochi, Lea M. Durham Delbridge, Mimi L. K. Tang, Leonard C. Harrison & Sarath Ranganathan
Sex Development Laboratory, Hudson Institute of Medical Research, Clayton, Australia
Vincent R. Harley & Yann W. Yap
Departments of Paediatrics and Community Health Sciences, The University of Calgary, Calgary, Canada
Deborah Dewey
Barwon Health, Geelong, Australia
Chloe J. Love, Peter Vuillermin, Fiona Collier & Lawrence Gray
Department of General Medicine, Royal Children’s Hospital, Parkville, Australia
David Burgner
Department of Pediatrics, Monash University, Clayton, Australia
Child Health Research Centre, The University of Queensland, Brisbane, Australia
Peter D. Sly
WHO Collaborating Centre for Children’s Health and Environment, Brisbane, Australia
Queensland Alliance for Environmental Health Sciences, The University of Queensland, Brisbane, Australia
Jochen F. Mueller
Monash Krongold Clinic, Faculty of Education, Monash University, Clayton, Australia
Nicole Rinehart
Centre for Developmental Psychiatry and Psychology, Monash University, Clayton, Australia
Bruce Tonge
School of BioSciences, Faculty of Science, The University of Melbourne, Parkville, Australia
Wah Chin Boon
Walter and Eliza Hall Institute, Parkville, Australia
Leonard C. Harrison
You can also search for this author in PubMed Google Scholar
the BIS Investigator Group
- , Toby Mansell
- , Martin O’Hely
- , David Burgner
- , Mimi L. K. Tang
- , Peter D. Sly
- , Richard Saffery
- , Jochen F. Mueller
- , Peter Vuillermin
- , Fiona Collier
- , Anne-Louise Ponsonby
- , Leonard C. Harrison
- , Sarath Ranganathan
- & Lawrence Gray
Contributions
Conceptualization—laboratory experiments: W.C.B., N.R., B.T., L.M.D.D. Laboratory experiments and analysis: W.C.B., K.V., S.D., H.K.C., J.C., F.C., CR, T.K., G.B.B., A.E.-B., S.E.H., N.T.T., K.B. Supervision of lab data collection: W.C.B., K.V., S.D., C.R. Laboratory statistical analysis: W.C.B., K.V., F.C., C.R., S.Th., V.H., Y.W.Y. Design and conduct of the Barwon Infant Study: C.S., A.-L.P., P.V., D.B., P.S., C.L., M.L.K.T., BIS Investigator Group. Design, conduct and analysis of the CCCEH-MN study: J.B.H., S.W., J.G. Design and conduct of BPA study measures in BIS: J.M., C.S., A.-.L.P. Design, conduct, and analysis of gene methylation studies: S.Ta., B.N., T.M., R.S., D.D., A.-L.P. Human studies statistical analysis: C.S., S.Th., A.-.L.P., S.Ta., K.V., M.O.H. Writing—reports and original draft: C.S., K.V., S.Th., S.Ta., A.-.L.P., W.C.B. Writing—editing: all authors. Results interpretation: all authors. Kara Britt did the laboratory experiment—estrogen pellet implantation.
Corresponding author
Correspondence to Wah Chin Boon .
Ethics declarations
Competing interests.
W.C.B. is a co-inventor on ‘Methods of treating neurodevelopmental diseases and disorders’, USA Patent No. US9925163B2, Australian Patent No. 2015271652. This has been licensed to Meizon Innovation Holdings. A.-L.P. is a scientific advisor and W.C.B. is a board member of the Meizon Innovation Holdings. The remaining authors declare no competing interests.
Peer review
Peer review information.
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information, peer review file, description of additional supplementary files, supplementary movie 1, supplementary movie 2, supplementary dataset 1, supplementary dataset 2, reporting summary, source data, source data, rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Cite this article.
Symeonides, C., Vacy, K., Thomson, S. et al. Male autism spectrum disorder is linked to brain aromatase disruption by prenatal BPA in multimodal investigations and 10HDA ameliorates the related mouse phenotype. Nat Commun 15 , 6367 (2024). https://doi.org/10.1038/s41467-024-48897-8
Download citation
Received : 01 December 2022
Accepted : 16 May 2024
Published : 07 August 2024
DOI : https://doi.org/10.1038/s41467-024-48897-8
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
By submitting a comment you agree to abide by our Terms and Community Guidelines . If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

- Open access
- Published: 14 August 2024
Mapping ultra-processed foods (UPFs) in India: a formative research study
- Suparna Ghosh-Jerath ORCID: orcid.org/0000-0002-2229-4455 1 na1 ,
- Neha Khandpur 2 na1 ,
- Gaurika Kumar 3 ,
- Sahiba Kohli 1 ,
- Meenu Singh 3 ,
- Inderdeep Kaur Bhamra 3 ,
- Fernanda H Marrocos-Leite 4 &
- K Srinath Reddy 3
BMC Public Health volume 24 , Article number: 2212 ( 2024 ) Cite this article
119 Accesses
Metrics details
Increased consumption of ultra-processed foods (UPFs) which have additives such as artificial colours, flavours and are usually high in salt, sugar, fats and specific preservatives, are associated with diet-related non-communicable diseases (NCDs). In India, there are no standard criteria for identifying UPFs using a classification system based on extent and purpose of industrial processing. Scientific literature on dietary intake of foods among Indian consumers classifies foods as unhealthy based on presence of excessive amounts of specific nutrients which makes it difficult to distinguish UPFs from other commercially available processed foods.
A literature review followed by an online grocery retailer scan for food label reading was conducted to map the types of UPFs in Indian food market and scrutinize their ingredient list for the presence of ultra-processed ingredients. All UPFs identified were randomly listed and then grouped into categories, followed by saliency analysis to understand preferred UPFs by consumers. Indian UPF categories were then finalized to inform a UPF screener.
A lack of application of a uniform definition for UPFs in India was observed; hence descriptors such as junk-foods , fast-foods , ready-to-eat foods , instant-foods , processed-foods , packaged-foods , high-fat-sugar-and-salt foods were used for denoting UPFs. After initial scanning of such foods reported in literature based on standard definition of UPFs, an online grocery retailer scan of food labels for 375 brands (atleast 3 brands for each food item) confirmed 81 food items as UPFs. A range of packaged traditional recipes were also found to have UPF ingredients. Twenty three categories of UPFs were then developed and subjected to saliency analysis. Breads, chips and sugar-sweetened beverages (e.g. sodas and cold-drinks) were the most preferred UPFs while frozen ready-to-eat/cook foods (e.g. chicken nuggets and frozen kebabs) were least preferred.
India needs to systematically apply a food classification system and define Indian food categories based on the level of industrial processing. Mapping of UPFs is the first step towards development of a quick screener that would generate UPF consumption data to inform clear policy guidelines and regulations around UPFs and address their impact on NCDs.
Peer Review reports
Non-communicable diseases (NCDs) are one of the leading causes of premature morbidity and mortality resulting in over 7 out of 10 deaths worldwide [ 1 ]. Mortality due to NCDs has been on the rise in India, increasing from 37.9% of all deaths in 1990 to 61.8% in 2016 [ 2 , 3 ]. Overweight/obesity have been identified as a contributing factor [ 4 ]. The recent national-level data shows an increase of 25% in the prevalence of overweight and obesity among Indian men and women over 14–15 years and 3% among children under five years [ 5 , 6 ]. Due to their thin fat phenotype, Indian infants and children, who comprise almost one quarter of the total population, are predisposed to obesity [ 7 , 8 ]. These risk factors are further amplified by changing food environments and behavioural variables such as tobacco, alcohol, drug use and low physical activity [ 9 ]. Exposure to unhealthy food environments in genetically predisposed children, along with other behavioural risk factors, increases their risk of developing obesity and diet-related non-communicable diseases (DR-NCDs) in the long term [ 10 ].
The rapidly changing food environment is characterized by diets transitioning from minimally-processed staple foods (such as pulses and whole cereals) high in vitamins, minerals and fibre to refined, processed and ultra-processed foods (UPFs) [ 11 ]. The Indian population is exposed to a wide variety of UPFs which are hyper-palatable, packaged, convenient, affordable and have a long shelf life, such as sugar-sweetened beverages, chips, biscuits and bread, and ready-to-eat/ ready-to-cook (RTC) meals [ 12 ]. The sales data of UPFs in India demonstrates an exponential increase, from USD 0.9 billion in 2006 to USD 37.9 billion in 2019 [ 13 ]. This growth indicates a notable expansion of these food products in the market, coupled with widespread advertising efforts that specifically target vulnerable populations, including children and youth [ 14 , 15 , 16 , 17 , 18 ]. Consumer demand for UPFs has increased due to higher disposable incomes, nuclear families, single-member households, and less availability of time for housework [ 19 , 20 ]. UPFs have penetrated the rural boundaries of the country and are likely making their way into households of diverse geographic and socio-economic attributes [ 21 , 22 ].
The Nova food classification system categorizes foods based on the purpose and the level of processing and includes four categories: (i) unprocessed/ minimally processed foods, (ii) processed culinary ingredients, (iii) processed foods, and (iv) ultra-processed foods [ 23 , 24 , 25 , 26 ]. UPFs are a category of food that undergo a series of industrial processes like extrusion and moulding, and have presence of classes of additives whose function is to make the final product palatable or more appealing, such as flavours, flavour enhancers, colours, emulsifiers, thickeners, sweeteners, etc. Although not unique to UPF, they also include additives that prolong the product duration and protect original properties or prevent proliferation of microorganisms [ 23 , 24 , 25 , 26 ]. In addition to this, several of these products are high in saturated fats or trans-fats, added sugars, and salt and low in dietary fibre, various micronutrients and other bioactive compounds [ 27 , 28 , 29 , 30 , 31 , 32 , 33 , 34 , 35 ].
Overconsumption of UPFs has been associated with higher body mass index (BMI), obesity, type-2 diabetes, hypertension, cardiovascular diseases, and certain types of cancers [ 36 , 37 , 38 ]. Given the diversity in UPFs, there is a need to systematically map the range of UPFs accessed by the Indian population. This is an important first step to understanding their potential role in contributing to the NCD burden in India and in developing strategies to encourage the substitution of the most frequently consumed UPFs to healthier alternatives. Identifying specific UPF categories could also help inform the development of dietary assessment instruments like food frequency questionnaires (FFQ) and screeners.
The present study aimed at: (i) mapping the specific categories of UPFs accessed and consumed in India, (ii) assessing the ingredient composition of these products, (iii) ranking the UPFs by consumer preference, and (iv) developing a list of categories of UPFs commonly consumed in India. For this, a secondary review of available literature complemented by an online grocery retailer scan and a saliency analysis were conducted between April 2021 and February 2022 (Fig. 1 ).
Flow chart briefly explaining the 3 steps of methodology
Step 1. The literature review was conducted to map and identify the various types of UPFs accessed, consumed, preferred and/ or purchased (as reported behaviours) in India. This review included published cross-sectional and observational research studies that used surveys, focus group discussions and interviews to elicit reported behaviours across different population groups and regions in India. International and national survey reports on UPF food intake and purchase among Indian population were also included in the review. Articles for review were identified from two electronic databases (NLM NCBI and Google Scholar). To ensure the search captured the diversity of UPFs, search terms included proxy descriptors identified in Indian policy documents [ 39 , 40 , 41 , 42 , 43 , 44 ], including: junk food* , fast food* , modern food* , westernized food* , ultra-processed food* , UPF* , convenience food* , ready-to-eat food* , ready-to-eat snack* , ready-to-cook food* , instant food* , frozen food* , canned food* , tinned food* , processed food* , packaged food* , high fat , sugar and salt food* and HFSS*. The literature search and data extraction was conducted by two authors (MS and GK).
To be eligible for the review, studies needed to: (i) include UPFs or their proxy descriptors, with examples of products, (ii) be conducted in either rural and/or urban areas of India, (iii) be published in the English language, between January 2012 and December 2022. This time frame was chosen to capture the high growth in UPFs sales during this decade [ 45 , 46 ]. Review articles, and publications that did not define the food category studied or did not cite any examples of foods were excluded.
Data from eligible articles were extracted in MS-Excel to record key variables on UPFs or their proxy descriptors with examples, location of the study (national/specific state), geographical area (urban/rural), sample size, sampling method, study participants’ age (years) and dietary data collection tools such as 24 h dietary recalls, FFQ, interviews and structured questionnaires (Additional file 1 ). A free list of UPF foods and beverages identified from the reviewed studies, was developed.
Step 2. An online grocery retailer scan for extracting detailed information on the UPFs identified in Step 1, was also conducted. The objective was to review and scrutinize the ingredient list provided in the food labels and to confirm that the food item qualified as UPFs. For this online scan, three researchers (GK, IKB, MS) reviewed the online grocery websites of the largest grocery retailers in India - Big Basket, Grofers, and Amazon [ 47 ]. Individual foods and beverages from Step 1, were checked for their ingredient composition and the presence of additives. This activity was guided by the FAO document ‘Ultra-processed foods, diet quality, and health using the Nova classification system’ [ 23 , 24 , 25 , 26 ] and the expertise of the co-authors (NK and FHML). Food items were specifically scrutinized for the use of additives (flavours, flavour enhancers, colours, emulsifiers, emulsifying salts, artificial sweeteners, thickeners, foaming, anti-foaming, bulking, carbonating, gelling and glazing agents), specific ingredients such as industrially derived sugars (fructose, invert sugar, maltodextrin, dextrose, lactose, high fructose corn syrup, fruit juice concentrates), modified oils (hydrogenated fats, interesterified fats), extracted proteins (hydrolysed proteins, soy protein isolate, gluten, casein, whey protein, mechanically separated meat) [ 23 , 24 , 25 , 26 , 48 ]. All food items were assessed for at least three different brands and if a majority of the items (2 out of 3) qualified as UPFs, the product category was confirmed as UPF. The free-list of UPFs identified from the literature and confirmed through label reading using the online grocery retailer scan were then categorized on the basis of the primary ingredient of their composition and/or functionality of the product. A 23 category UPF list was developed at the end of Step 2.
Step 3. The confirmed UPF categories (23 categories), were then subjected to a saliency analysis, conducted by two authors (GK, MS). Saliency is a statistical accounting of items for rank and frequency of mention, across all respondents within a given domain. For example, the colour chosen most often from a free list of ten colours by a study population is referred to as the most salient [ 49 ]. The saliency test indicators included the commonly accessed, consumed, preferred and/or purchased (collectively referred to as ‘preferred’ in this paper to identify the common UPF categories accessed by the Indian consumers). These categories were limited to the food items that were confirmed as UPFs in Step 2. For example, if a study used “junk food” as a descriptor of UPFs and included freshly prepared savouries like “samosa/kachori” along with chips and soft drinks, we included data for only chips and soft drinks for the purpose of saliency analysis. The UPFs were then sorted from the most to the least preferred UPFs (Additional file 2 ). The steps and formulas [ 49 ] used to calculate composite salience scores for each UPF category have been illustrated in Fig. 2 . The UPF categories were classified per consumer preference, to the composite salience score cut-offs, defined after dividing the distribution of the composite salience scores obtained into tertiles as follows: (i) ≥ 0.61 as frequently preferred, (ii) 0.61 − 0.51 as infrequently preferred, and (iii) < 0.51 as rarely preferred UPFs.
Saliency analysis method for free-listed UPF categories
The literature search and study selection process of Step 1 is illustrated in Fig. 3 . A total of 23 research articles that matched the inclusion criteria were included in the final review. An overview of the extracted variables is provided in Additional File 1 . These studies were conducted in both rural (5 out of 23) and urban areas (17 out of 23), in different regions of the country, among a diverse population aged between 9 and 69 years (Table 1 ). Table 2 provides the outcome of the literature review with proxy descriptors along with the food items listed under them. These foods were verified as UPFs and non-UPFs.
Flow diagram reporting the screening and selection of studies reporting consumption and availability of UPFs in India
The online grocery retailer scan, label readings of 375 packaged foods were completed (atleast 3 brands per product) and 81 of those food products qualified as UPFs. Several of the packaged Indian traditional foods and snacks such as bottled and packaged pickles, namkeens (cereal and pulse-based extruded snacks), papads , frozen non-vegetarian meals and snacks, and frozen RTE meals (like rajmah curry and rice, biryani , dal makhni , etc.) had UPF ingredients and additives in their formulation that qualified them as UPFs (Table 3 ). Food products such as RTE breakfast cereals (e.g. poha , upma , etc.), RTE Indian curries (e.g. paneer makhani , butter chicken, etc.), Indian RTE bread (e.g. thepla , paratha , etc.), RTC mixes (e.g. idli mix, dal vada mix, etc.) also qualified as UPFs. However, some RTE traditional meals such as RTE biryani , RTE rajmah curry with rice, RTE kadhi pakoda with rice were not categorized as UPFs as these did not include UPF ingredients.
Consumer preferences for the confirmed UPF food categories identified above, were assessed using saliency scores. Table 4 lists these categories and shows the order of preference based on the saliency scores. The last column in the table indicates ‘frequently’, ‘infrequently’ or ‘rarely’ preferred UPFs by consumers in India. The frequently preferred UPFs were breads, chips and other extruded snacks (such as potato chips, cheese balls, puff corns, etc.) and sugar-sweetened beverages (such as cold drinks, diet coke, sodas, and energy drinks. The three rarely preferred UPFs were margarine and frozen/ packaged vegetarian and non-vegetarian snacks and meals (such as stuffed/plain parantha , naan , palak paneer , rajma , cutlets, fish/seafood snacks, salami, and sausages).
The present study aimed to identify the specific categories of UPFs accessed in India and rank them by consumer preference using a literature review, an online grocery retailer scan and saliency analysis. We found 23 categories of UPFs accessed by Indian consumers. After analysing the ingredient list of UPFs, we found that product formulation of several traditional Indian foods has transitioned from being processed to ultra-processed category with the use of industrially processed ingredients and presence of additives such as artificial colours, flavour enhancers, anti-caking agents. These ultra-processed versions of traditional foods even though have similar nutrient composition to home-prepared meals, are increasingly consumed, displacing home-cooked meals, and substituting staples. While the health effects of this displacement from minimally processed food ingredients to UPFs is an area of on-going research, we have growing evidence that UPF dietary patterns are linked to poor health outcomes [ 23 , 69 ]. It is crucial to track reformulation of traditional recipes to ultra-processed convenience foods especially since traditional meals are thought to be healthier [ 70 ]. The increasing market of ultra-processed traditional Indian recipes with poor nutritional profile needs more scrutiny and research.
Saliency analysis identified the preferred UPFs among the Indian population with breads, chips and sugar-sweetened beverages being the most preferred UPFs and frozen non-vegetarian snacks being the least preferred. This finding is consistent with the sales trends reported by Euromonitor International in 2020, which has also highlighted a substantial contribution of similar categories of packaged foods, such as bakery items, biscuits, packaged dairy products, savory snacks, and sauces and condiments [ 46 ]. Further, saliency analysis also indicates the preference of Indian consumers towards UPFs such as fruit-based preserves, cookies and biscuits, Indian sweet mixes, sauces and pickles, instant noodles/soups/ pasta and savoury puff rolls. Studies from other low and middle-income countries (LMICs) demonstrates similar trends in preference (consumption of UPFs and contribution to percentage of total calories) towards packaged confectioneries, savoury snacks, deep-fried foods, biscuits, candy/ chocolate, savoury snacks, canned red and luncheon meats, pre-fried French fries, mayonnaise, ketchup, fast-food such as sandwiches and pizzas, chips and salty snacks (including tortillas and pretzels), sweets and sweetened beverages and sausages (including canned) [ 71 , 72 ].
Our results also suggest a benefit of utilizing a classification system based on processing. Currently several UPFs are being captured by proxy descriptors like junk foods, fast foods, convenience foods, instant foods, packaged foods, etc. This limits comparability with other studies, monitoring the preference for and consumption of these products by the population, developing targeted interventions, tracking product reformulation and other regulatory measures to control exposure of these foods to vulnerable age groups through food advertising, etc. [ 73 ]. Using UPFs more consistently in studies reporting unhealthy food consumption pattern in India will help with global comparisons and in also elucidating the health effects of these foods. Additionally, as per the packaged food sales data from 2015-19, the Indian UPF market is slowly expanding with increasing sales of RTE meals, savoury snacks, processed fruits, vegetables, meats and other packaged foods [ 46 ]. The Nova food classification system can serve this purpose and may be explored as an option for categorization of foods by regulatory authorities. This classification system is used to assess dietary patterns in several high and middle-income countries [ 23 , 70 ]. Food based dietary guidelines of several countries such as Brazil, Uruguay, Ecuador, Peru and Israel have utilized Nova classification system to inform their dietary recommendations [ 74 , 75 , 76 , 77 , 78 ].
The present paper identified only a limited number of Indian studies which were primarily reported from 2 geographical regions. More such surveys on the consumption of UPFs are desirable to identify common regional UPFs. In the Indian context, several UPFs are indigenously produced by local retailers apart from the huge market share of nationally known branded UPFs [ 79 ]. These locally accessible UPFs have greater penetration into the local markets.
The categories of UPFs in India developed in the present study after due validation can be developed into a UPF consumption screener. This tool can be used for monitoring the UPF consumption in India and can address critical gap in scientific literature. This information on quantitative estimate of UPF consumption among Indian population can be useful for assessing impact of UPF consumption on increasing burden of NCDs in India.
This study is one of the first attempts to explore the types of UPFs in the Indian food market, identify the types of packaged traditional recipes that have been converted to UPFs, and map their saliency.
Study limitations
Studies reviewed were majorly from South India and largely represented the urban population, hence the results cannot be extended to the rural population. The study could only conduct saliency mapping of preferred foods without quantity of intake of UPFs and their contribution to total day’s energy intake. We could not explore traditional variants of UPFs that may be sold in the local unregulated markets.
Conclusions
India needs to develop a food classification system while systematically defining food categories based on level of processing. This should be followed by an assessment of the extent of UPFs consumption in India. The mapping of the UPFs in India reported in this paper provides the first step in developing a quick screener that systematically lists all the UPF categories. The data generated on consumption of UPFs using the screener is likely to inform policies on regulating the Indian UPFs market, undertake consumer education initiatives and create nutrition literacy around UPFs and thus contain their indiscriminate consumption. This may address the impact of UPF consumption on increasing burden of NCDs in India. There is an urgent need for strengthening the food regulatory environment to check the infiltration of several unhealthy UPFs in the Indian food market.
Data availability
No new data was created or analyzed under the literature review part of the study. The datasets used as part of a particular component is available from the corresponding author on reasonable request.
Abbreviations
- Ultra-processed foods
Non-communicable diseases
Diet-related non-communicable diseases
Food frequency questionnaires
High fat sugar salt
Low and middle-income countries
Ready-to-eat
Ready-to-cook
World Health Organization. Non-Communicable Diseases Country Profiles 2018. https://apps.who.int/iris/handle/10665/274512 . [Cited 18 January 2023].
World Health Organization. Non-Communicable Diseases Progress Monitor 2022. https://apps.who.int/iris/handle/10665/353048 . [Cited 18 January 2023].
Indian Council of Medical Research (ICMR), Public Health Foundation of India (PHFI) and Institute for Health Metrics and Evaluation (IHME). India: Health of the Nation’s States-The Indian State-level Disease Burden Initiative. New Delhi: ICMR, PHFI, and IHME; 2017). https://phfi.org/downloads/171110_India_Health_of_Nation_states_Report_2017.pdf [Cited 20 January 2023].
Alwan A. Global status report on non-communicable diseases 2010. World Health Organization, 2011. https://apps.who.int/iris/bitstream/handle/10665/44579/9789240686458_eng.pdf . [Cited 8 May 2023].
International Institute for Population Sciences (IIPS) and ICF. National Family Health Survey (NFHS-5), India, 2019-21: Mizoram. Mumbai: IIPS. 2021. https://pdf.usaid.gov/pdf_docs/PA00ZBN4.pdf . [Cited 18 January 2023].
Sethi V, Lahiri A, Bhanot A, Kumar A, Chopra M, Mishra R, Alambusha R, Agrawal P, Johnston R, de Wagt A. Adolescents, diets and Nutrition: growing well in a changing World, the Comprehensive National Nutrition Survey. Thematic Rep, (1), 2019.
D’Angelo S, Yajnik CS, Kumaran K, Joglekar C, Lubree H, Crozier SR et al. Body size and body composition: a comparison of children in India and the UK through infancy and early childhood. J Epidemiol Community Health 2015 July 16;69(12):1147.
India - Sample Registration System (SRS)-Statistical Report. 2020. https://censusindia.gov.in/nada/index.php/catalog/44376 . [Cited 18 January 2023].
Dandona L, Dandona R, Kumar GA, Shukla DK, Paul VK, Balakrishnan K, et al. Nations within a nation: variations in epidemiological transition across the States of India, 1990–2016 in the global burden of Disease Study. Lancet. 2017;390(10111):2437–60.
Article Google Scholar
Tak M, Law C, Green R, Shankar B, Cornelsen L. Processed foods purchase profiles in urban India in 2013 and 2016: a cluster and multivariate analysis. BMJ open. 2022;12(10):e062254.
Article PubMed PubMed Central Google Scholar
Baker P, Friel S. Processed foods and the nutrition transition: evidence from Asia. Obes Rev. 2014;15(7):564–77.
Article CAS PubMed Google Scholar
Dunford EK, Farrand C, Huffman MD, Raj TS, Shahid M, Ni Mhurchu C, Neal B, Johnson C. Availability, healthiness, and price of packaged and unpackaged foods in India: a cross-sectional study. Nutr Health. 2022;28(4):571–9.
Article PubMed Google Scholar
The growth of ultra-processed foods in India: an analysis of trends, issues and policy recommendations. New Delhi: World Health Organization, Country Office for India; 2023. https://www.who.int/publications/i/item/9789290210672
Bassi S, Bahl D, Arora M, Tullu FT, Dudeja S, Gupta R. Food environment in and around schools and colleges of Delhi and National Capital Region (NCR) in India. BMC Public Health. 2021;21(1):1–3.
Harris J, Heard A, Schwartz M. Older but still vulnerable: all children need protection from unhealthy food marketing. Yale Rudd Center for Food Policy & Obesity. 2014; www.uconnruddcenter.org/files/Pdfs/Protecting_Older_Children_3_14.pdf . [Cited 24 December 2022].
Euromonitor International. (2019). Euromonitor International; London. Passport Global Market Information Database. https://www.euromonitor.com/
Pandav C, Smith Taillie L, Miles DR, Hollingsworth BA, Popkin BM. The WHO South-East Asia region nutrient profile model is quite appropriate for India: an exploration of 31,516 food products. Nutrients. 2021;13(8):2799.
The growth of ultra-processed foods in India: an analysis of trends, issues and policy recommendations. New Delhi: World Health Organization, Country Office for India; 2023.
Monteiro CA, Moubarac JC, Cannon G, Ng SW, Popkin B. Ultra-processed products are becoming dominant in the global food system. Obes Rev. 2013;14:21–8.
Magalhães V, Severo M, Correia D, Torres D, de Miranda RC, Rauber F, Levy R, Rodrigues S, Lopes C. Associated factors to the consumption of ultra-processed foods and its relation with dietary sources in Portugal. J Nutritional Sci. 2021;10.
Prakash J. Consumption trends of processed foods among rural population selected from South India. Int J Food Nutr Sci. 2015;2(6):1–6.
Google Scholar
Gupta A, Kapil U, Singh G. Consumption of junk foods by school-aged children in rural Himachal Pradesh, India. Indian J Public Health. 2018;62(1):65.
Monteiro CA, Cannon G, Lawrence M, da Costa Louzada ML, Pereira Machado P. Ultra-processed foods, diet quality, and health using the NOVA classification system. Rome: FAO; 2019. p. 48.
Monteiro CA, Cannon G, Levy RB, Moubarac JC, Louzada ML, Rauber F, Khandpur N, Cediel G, Neri D, Martinez-Steele E, Baraldi LG. Ultra-processed foods: what they are and how to identify them. Public Health Nutr. 2019;22(5):936–41.
Monteiro CA, Cannon G, Levy R, Moubarac J-C, Jaime P, Martins AP, et al. NOVA: the star shines bright. World Nutr. 2016;7(1–3):28–38.
Monteiro CA, Levy RB, Claro RM, Castro IRR, Cannon G. A new classification of foods based on the extent and purpose of their processing. Cad Saude Publica. 2010;26:2039–49. https://doi.org/10.1590/S0102-311X2010001100005 .
Moubarac JC, Batal M, Louzada ML, Steele EM, Monteiro CA. Consumption of ultra-processed foods predicts diet quality in Canada. Appetite. 2017;108:512–20.
Rauber F, Louzada ML, Steele EM, Millett C, Monteiro CA, Levy RB. Ultra-processed food consumption and chronic non-communicable diseases-related dietary nutrient profile in the UK (2008–2014). Nutrients. 2018;10(5):587.
da Costa Louzada ML, Ricardo CZ, Steele EM, Levy RB, Cannon G, Monteiro CA. The share of ultra-processed foods determines the overall nutritional quality of diets in Brazil. Public Health Nutr. 2018;21(1):94–102.
Louzada ML, Martins AP, Canella DS, Baraldi LG, Levy RB, Claro RM, Moubarac JC, Cannon G, Monteiro CA. Impact of ultra-processed foods on micronutrient content in the Brazilian diet. Rev Saude Publica. 2015;49.
Louzada ML, Martins AP, Canella DS, Baraldi LG, Levy RB, Claro RM, Moubarac JC, Cannon G, Monteiro CA. Ultra-processed foods and the nutritional dietary profile in Brazil. Rev Saude Publica. 2015;49.
Monteiro CA, Levy RB, Claro RM, de Castro IR, Cannon G. Increasing consumption of ultra-processed foods and likely impact on human health: evidence from Brazil. Public Health Nutr. 2011;14(1):5–13.
Moubarac JC, Martins AP, Claro RM, Levy RB, Cannon G, Monteiro CA. Consumption of ultra-processed foods and likely impact on human health. Evidence from Canada. Public Health Nutr. 2013;16(12):2240–8.
Poti JM, Mendez MA, Ng SW, Popkin BM. Is the degree of food processing and convenience linked with the nutritional quality of foods purchased by US households? Am J Clin Nutr. 2015;101(6):1251–62.
Article CAS PubMed PubMed Central Google Scholar
Luiten CM, Steenhuis IH, Eyles H, Mhurchu CN, Waterlander WE. Ultra-processed foods have the worst nutrient profile, yet they are the most available packaged products in a sample of New Zealand supermarkets. Public Health Nutr. 2016;19(3):530–8.
Beslay M, Srour B, Méjean C, Allès B, Fiolet T, Debras C, Chazelas E, Deschasaux M, Wendeu-Foyet MG, Hercberg S, Galan P. Ultra-processed food intake in association with BMI change and risk of overweight and obesity: a prospective analysis of the French NutriNet-Santé cohort. PLoS Med. 2020;17(8):e1003256.
Levy RB, Rauber F, Chang K, Louzada ML, Monteiro CA, Millett C, Vamos EP. Ultra-processed food consumption and type 2 diabetes incidence: a prospective cohort study. Clin Nutr. 2021;40(5):3608–14.
de Miranda RC, Rauber F, de Moraes MM, Afonso C, Santos C, Rodrigues S, Levy RB. Consumption of ultra-processed foods and non-communicable disease-related nutrient profile in Portuguese adults and elderly (2015–2016): the UPPER project. Br J Nutr. 2021;125(10):1177–87.
Food Safety and Standards. (Safe food and balanced diets for School Children) Regulations 2020. https://www.fssai.gov.in/upload/uploadfiles/files/Gazette_Notification_Safe_Food_Children_07_09_2020.pdf . [Cited 12 May, 2023].
Report of Working Group on Addressing Consumption of Foods High in Fat. Salt and Sugar (HFSS) and Promotion of Healthy Snacks in Schools of India. Working Group Constituted by Ministry of Women and Child Development Government of India 2015. https://www.nipccd.nic.in/file/reports/hfss.pdf . [Cited 12 May, 2023].
What India Eats ICMR-NIN. 2020. https://www.millets.res.in/pdf/what_india_eats.pdf . [Cited 12 May, 2023].
National Programme for Prevention and Control of Cancer, Diabetes, Cardiovascular Diseases and Stroke (NPCDCS). Directorate General of Health Services, Ministry of Health and Family Welfare, Government of India. Handbook for Counsellors Reducing Risk Factors for Non-Communicable Disease. 2017 Sep. https://main.mohfw.gov.in/sites/default/files/Handbook%20for%20Counselors%20-%20Reducing%20Risk%20Factors%20for%20NCDs_1.pdf . [Cited 12 May,2023].
Recommended Dietary Allowances and Estimated Average Requirements Nutrient Requirements for Indians. A Report of the Expert Group - Indian Council of Medical Research National Institute of Nutrition 2020. https://www.nin.res.in/RDA_Full_Report_2020.html . [Cited 12 May, 2023].
National Multi-sectoral Action Plan for Prevention and Control of Common. NCDs 2017–2022. https://main.mohfw.gov.in/sites/default/files/National%20Multisectoral%20Action%20Plan%20%28NMAP%29%20for%20Prevention%20and%20Control%20of%20Common%20NCDs%20%282017-22%29_1.pdf ,. [Cited 12 May, 2023].
Baker P, Machado P, Santos T, Sievert K, Backholer K, Hadjikakou M, Russell C, Huse O, Bell C, Scrinis G, Worsley A. Ultra-processed foods and the nutrition transition: global, regional and national trends, food systems transformations and political economy drivers. Obes Rev. 2020;21(12):e13126.
Packaged Food in India. Euromonitor Int 2020. https://www.euromonitor.com/
India Online Grocery Market Size. Report 2021–2028. Grand View Research. https://www.grandviewresearch.com/industry-analysis/india-online-grocery-market . [Cited 12 May, 2023].
Food Safety and Standards (Food Products Standards and Food Additives) Regulations. 2011. https://www.fssai.gov.in/cms/food-safety-and-standards-regulations.php . [Cited 8 May, 2023].
Quinlan M. Considerations for collecting free-lists in the field: examples from ethobotany. Field Methods. 2005;17(3):219–34.
Jain A, Mathur P. Intake of ultra-processed foods among adolescents from low-and middle-income families in Delhi. Indian Pediatr. 2020;57(8):712–4.
National Non-communicable Disease Monitoring Survey (NNMS 2017-18), Bengaluru ICMR-NCDIR. India. 2021. Available from: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/viewer.html?Pdfurl = https%3A%2F%2Fwww.ncdirindia.org%2Fnnms%2Fresources%2fchapter_4_2_3.pdf&clen = 181451&chunk = true. [Cited 18 Jan, 2023]
Law C, Green R, Kadiyala S, Shankar B, Knai C, Brown KA, Dangour AD, Cornelsen L. Purchase trends of processed foods and beverages in urban India. Global Food Secur. 2019;23:191.
Goel S, Kaur T, Gupta M. Increasing proclivity for junk food among overweight adolescent girls in district Kurukshetra, India. Int Res J Biol Sci. 2013;16:17.
Bhavani V, Prabhavathy Devi N. Junk and Sink: A Comparative Study on Junk Food Intake among Students of India’. Shanlax Int J Arts Sci Humanit. 2020:13–8.
Naveenkumar D, Parameshwari S. A study on junk food consumption and its Effect on adolescents in Thiruvannamalai District, Tamil Nadu, India. Adalya J August. 2019;8(8):781–93. [ISSN 1301–2746].
Hiregoudar V, Suresh CM, Singode C, Raghavendra B. Proportion, patterns, and determinants of junk food consumption among adolescent students. Annals Community Health. 2021;9(2):46.
Kumari R, Kumari M. Quality aspects of fast foods and their consumption pattern among teenagers of rural-urban region of Sabour block in Bhagalpur district of India. The Pharma Innovation Journal. 2020. 2020;9(4):96–102.
Amin T, Choudhary N, Jabeen AN. Study of fast food consumption pattern in India in Children aged 16–20 years. Intl J Food Ferment Technol. 2017;7(1):1–8.
Singh M, Mishra S. Fast food consumption pattern and obesity among school going (9–13 year) in Lucknow District. Int J Sci Res. 2014;3(6):1672–4.
Kotecha PV, Patel SV, Baxi RK, Mazumdar VS, Shobha M, Mehta KG, Mansi D, Ekta M. Dietary pattern of school going adolescents in urban Baroda, India. J Health Popul Nutr. 2013;31(4):490.
CAS PubMed PubMed Central Google Scholar
Joseph N, Nelliyanil M, Rai S, YP RB, Kotian SM, Ghosh T, Singh M. Fast food consumption pattern and its association with overweight among high school boys in Mangalore city of southern India. J Clin Diagn Research: JCDR. 2015;9(5):LC13.
Prabhu NB. Examining fast-food consumption behaviour of students in Manipal University, India. Afr J Hospitality Tourism Leisure. 2015;4(2):621–30.
Khongrangjem T, Dsouza SM, Prabhu P, Dhange VB, Pari V, Ahirwar SK, Sumit K. A study to assess the knowledge and practice of fast food consumption among pre-university students in Udupi Taluk, Karnataka, India. Clin Epidemiol Global Health. 2018;6(4):172–5.
Landge JA, Khadkikar GD. Lifestyle and nutritional status of late adolescent in an urban area of Western Maharashtra: cross sectional study. Int J Community Med Public Health. 2020;7(8):3027.
Iyer ML, Kumar NP. A study of Breakfast habits of Urban Indian consumers. Research Journals of Economics and Business Study. Res J Econ Bus Stud. 2014;3(8):107–16. [ISSN: 2251 – 1555].
Rathi N, Riddell L, Worsley A. Food consumption patterns of adolescents aged 14–16 years in Kolkata, India. Nutr J. 2017;16(1):1–2.
Vasan M. Consumers’ preference and consumption towards instant Food products. Think India J. 2019;22(14):8333–7.
Harrell M, Medina J, Greene-Cramer B, Sharma SV, Arora M. Understanding eating behaviors of New Delhi’s Youth. J Appl Res Child. 2015;6(2):8.
Mediratta S, Ghosh S, Mathur P. Intake of ultra-processed food, dietary diversity and the risk of nutritional inadequacy among adults in India. Public Health Nutr. 2023;26(12):2849–58.
Monteiro CA, Cannon G, Moubarac JC, Levy RB, Louzada MLC, Jaime PC. The UN Decade of Nutrition, the NOVA food classification and the trouble with ultra-processing. Public Health Nutr. 2017;21(1):5–17.
Islam MR, Rahman SM, Rahman MM, Pervin J, Rahman A, Ekström EC. Gender and socio-economic stratification of ultra-processed and deep-fried food consumption among rural adolescents: a cross-sectional study from Bangladesh. PLoS ONE. 2022;17(7):e0272275.
Pries AM, Filteau S, Ferguson EL. Snack food and beverage consumption and young child nutrition in low-and middle‐income countries: a systematic review. Matern Child Nutr. 2019;15:e12729.
Costa CD, Faria FR, Gabe KT, Sattamini IF, Khandpur N, Leite FH, Steele EM, Louzada ML, Levy RB, Monteiro CA. Nova score for the consumption of ultra-processed foods: description and performance evaluation in Brazil. Rev Saúde Pública. 2021;55:13.
Monteiro CA, Cannon G, Moubarac JC, Martins AP, Martins CA, Garzillo J, Canella DS, Baraldi LG, Barciotte M, Louzada ML, Levy RB, Claro RM, Jaime PC. Dietary guidelines to nourish humanity and the planet in the twenty-first century. A blueprint from Brazil. Public Health Nutr. 2015;18(13):2311–22. Epub 2015 Jul 24. PMID: 26205679; PMCID: PMC10271430.
Ministerio, de Salud. Guía Alimentaria para la Población Uruguay. 2016. https://www.gub.uy/ministerio-salud-publica/sites/ministerio-salud-publica/files/documentos/campanas/MSP_GUIA_ALIMENTARIA_POBLACION.pdf
Ministerio de Salud Pública del Ecuador y la Organización de las Naciones Unidas para la Alimentación y la Agricultura. Documento Técnico De las Guías Alimentarias Basadas en Alimentos (GABA) Del Ecuador. Quito. 2020. https://doi.org/10.4060/ca9928es .
Ministerio de Salud Instituto Nacional de Salud y Centro Nacional de Alimentación y Nutrición. Guías Alimentarias para la Población Peruana. 2020. https://repositorio.ins.gob.pe/bitstream/handle/20.500.14196/1247/Gu%c3%ada-alimen21.pdf?sequence=1&isAllowed=y
Nutritional Recommendations: The Israeli Ministry of Health. 2019. https://www.health.gov.il/PublicationsFiles/dietary%20guidelines%20EN.pdf
Downs SM, Thow AM, Ghosh-Jerath S, Leeder SR. Developing interventions to reduce consumption of unhealthy fat in the food retail environment: a case study of India. J Hunger Environ Nutr. 2014;9(2):210–29.
Download references
Acknowledgements
We would like to appreciate the contribution of Dr. Shukrani Shinde for supporting the study team during the literature search and online grocery retailer scan.
This work is funded through the Innovative Methods and Metrics for Agriculture and Nutrition Action (IMMANA) programme (Grant IMMANA 3.06), led by the London School of Hygiene & Tropical Medicine (LSHTM). IMMANA is co-funded with UK Aid from the UK government and by the Bill & Melinda Gates Foundation. This work was supported, in part, by the Bill & Melinda Gates Foundation [INV-002962 / OPP1211308]. Under the grant conditions of the Foundation, a Creative Commons Attribution 4.0 Generic License has already been assigned to the Author Accepted Manuscript version that might arise from this submission.
Author information
Suparna Ghosh-Jerath and Neha Khandpur contributed equally to this work.
Authors and Affiliations
The George Institute for Global Health, 308, Third Floor, Elegance Tower, Plot No. 8, Jasola District Centre, New Delhi, Delhi, 110025, India
Suparna Ghosh-Jerath & Sahiba Kohli
Wageningen University, Wageningen, The Netherlands
Neha Khandpur
Public Health Foundation of India, New Delhi, India
Gaurika Kumar, Meenu Singh, Inderdeep Kaur Bhamra & K Srinath Reddy
Center for Epidemiological Research in Nutrition and Health, Faculty of Public Health, University of Sao Paulo, Sao Paulo, Brazil
Fernanda H Marrocos-Leite
You can also search for this author in PubMed Google Scholar
Contributions
The study was conceptualized by KSR, SGJ, NK and FHML. The literature review and online grocery retailer scan were conducted by MS, GK, IKB. The first draft of the manuscript was prepared by MS, GK, IKB, SK and SGJ. The manuscript was critiqued and edited by SGJ, NK, FHML and KSR. SGJ had primary responsibility for final content; and all authors read and approved the final manuscript.
Corresponding author
Correspondence to Suparna Ghosh-Jerath .
Ethics declarations
Ethics approval and consent to participate.
This study was conducted according to guidelines laid down in the Declaration of Helsinki, and all procedures involving humans were approved by the Institutional Ethics Committee at the Public Health Foundation of India, and the ethics committee of University of Sao Paulo. The current manuscript, however, reports findings from an exhaustive literature review and online grocery retailer scan for which informed consent process is not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary material 2, rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Cite this article.
Ghosh-Jerath, S., Khandpur, N., Kumar, G. et al. Mapping ultra-processed foods (UPFs) in India: a formative research study. BMC Public Health 24 , 2212 (2024). https://doi.org/10.1186/s12889-024-19624-1
Download citation
Received : 04 July 2023
Accepted : 29 July 2024
Published : 14 August 2024
DOI : https://doi.org/10.1186/s12889-024-19624-1
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Nova food classification
- UPF categories
- Online grocery retailer scan
- Tool adaptation
- Nova-UPF screener
BMC Public Health
ISSN: 1471-2458
- General enquiries: [email protected]
COMMENTS
Tips on how to write a review of related literature in research. Given that you will probably need to produce a number of these at some point, here are a few general tips on how to write an effective review of related literature 2. Define your topic, audience, and purpose: You will be spending a lot of time with this review, so choose a topic ...
A literature review is a survey of scholarly sources (such as books, journal articles, and theses) related to a specific topic or research question. It is often written as part of a thesis, dissertation , or research paper , in order to situate your work in relation to existing knowledge.
A literature review is meant to analyze the scholarly literature, make connections across writings and identify strengths, weaknesses, trends, and missing conversations. A literature review should address different aspects of a topic as it relates to your research question. A literature review goes beyond a description or summary of the ...
A literature review may consist of simply a summary of key sources, but in the social sciences, a literature review usually has an organizational pattern and combines both summary and synthesis, often within specific conceptual categories.A summary is a recap of the important information of the source, but a synthesis is a re-organization, or a reshuffling, of that information in a way that ...
A literature review is an integrated analysis-- not just a summary-- of scholarly writings and other relevant evidence related directly to your research question.That is, it represents a synthesis of the evidence that provides background information on your topic and shows a association between the evidence and your research question.
Literature Review is a comprehensive survey of the works published in a particular field of study or line of research, usually over a specific period of time, in the form of an in-depth, critical bibliographic essay or annotated list in which attention is drawn to the most significant works.. Also, we can define a literature review as the collected body of scholarly works related to a topic:
Closely related to the semi-structured review approach is the integrative or critical review approach. ... review, an integrative review usually has a different purpose, with the aim to assess, critique, and synthesize the literature on a research topic in a way that enables new theoretical frameworks and perspectives to emerge (Torraco, 2005 ...
What kinds of literature reviews are written? Narrative review: The purpose of this type of review is to describe the current state of the research on a specific topic/research and to offer a critical analysis of the literature reviewed. Studies are grouped by research/theoretical categories, and themes and trends, strengths and weakness, and gaps are identified.
The broader scholarly literature should be related to the chosen main topic for the LR (how to develop the literature review section). The LR can cover the literature from one or more disciplines, depending on its scope, but it should always offer a new perspective. ... Ideas and theories in the field are related to research methodologies .
A literature review is a written work that: Compiles significant research published on a topic by accredited scholars and researchers; Surveys scholarly articles, books, dissertations, conference proceedings, and other sources; Examines contrasting perspectives, theoretical approaches, methodologies, findings, results, conclusions.
A literature or narrative review is a comprehensive review and analysis of the published literature on a specific topic or research question. The literature that is reviewed contains: books, articles, academic articles, conference proceedings, association papers, and dissertations. It contains the most pertinent studies and points to important ...
2. MOTIVATE YOUR RESEARCH in addition to providing useful information about your topic, your literature review must tell a story about how your project relates to existing literature. popular literature review narratives include: ¡ plugging a gap / filling a hole within an incomplete literature ¡ building a bridge between two "siloed" literatures, putting literatures "in conversation"
A literature review is a comprehensive summary of previous research on a topic. The literature review surveys scholarly articles, books, and other sources relevant to a particular area of research. The review should enumerate, describe, summarize, objectively evaluate and clarify this previous research. It should give a theoretical base for the ...
A literature review is a critical analysis and synthesis of existing research on a particular topic. It provides an overview of the current state of knowledge, identifies gaps, and highlights key findings in the literature. 1 The purpose of a literature review is to situate your own research within the context of existing scholarship ...
A review of related literature (RRL) is a part of the research report that examines significant studies, theories, and concepts published in scholarly sources on a particular topic. An RRL includes 3 main components: A short overview and critique of the previous research.
Literature reviews are in great demand in most scientific fields. Their need stems from the ever-increasing output of scientific publications .For example, compared to 1991, in 2008 three, eight, and forty times more papers were indexed in Web of Science on malaria, obesity, and biodiversity, respectively .Given such mountains of papers, scientists cannot be expected to examine in detail every ...
Types of Literature Review are as follows: Narrative literature review: This type of review involves a comprehensive summary and critical analysis of the available literature on a particular topic or research question. It is often used as an introductory section of a research paper. Systematic literature review: This is a rigorous and ...
A review of literature is a classification and evaluation of what accredited scholars and. researchers have written on a topic, organized according to a guiding concept such as a research ...
The literature review provides a way for the novice researcher to convince the proposal the reviewers that she is knowledgeable about the related research and the "intellectual traditions" that support the proposed study. The literature review provides the researcher with an opportunity to identify any gaps that may exist in the body of ...
A comparison/contrast of the choices different authors or characters make in a work. A reading of a work based on an outside philosophical perspective (Ex. how would a Freudian read Hamlet ?) A study of the sources or historical events that occasioned a particular work (Ex. comparing G.B. Shaw's Pygmalion with the original Greek myth of ...
Join the 250,000+ researchers, students, and professionals using Litmaps to accelerate their literature review. Find the right papers faster. Get started for free! ... Using Litmaps for my research papers has significantly improved my workflow. Typically, I start with a single paper related to my topic. ... University of Applied Sciences ...
Writing the Literature Review Barry Mauer and John Venecek. We discuss the following topics on this page: The Literature Review; ... The aim of a scoping review is to provide a comprehensive overview or map of the published research or evidence related to a research question. This might be considered a prelude to a systematic review that would ...
Finding "The Literature" Organizing/Writing; APA Style This link opens in a new window; Chicago: Notes Bibliography This link opens in a new window; MLA Style This link opens in a new window; Sample Literature Reviews. Sample Lit Reviews from Communication Arts; Have an exemplary literature review? Get Help!
R.I.E., Bhopal. [email protected]. Introduction. "Review of related Literature" is mostly known as the second chapter in almost every. empirical thesis. But it is the very first step to ...
A review of related literature (RRL) puts your research in the context of what is currently known about the topic. Accordingly, it is a good way to highlight the knowledge gaps and what your study offers in terms of novelty and significance. Your question on outlining the subtopics is a pertinent one. However, outlining subtopics would heavily ...
As already indicated by others, a literature review should relate to your research question, so have a look what the relation is between your approach and that of others, where it differs, adds ...
Looking in the reference section of a literature or systematic review can be a good place to find original research studies. Below is a screenshot of the abstract of the article The Effect of Dietary Glycaemic Index on Glycaemia in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials , with words ...
Generating knowledge graphs through text mining of catalysis research related literature†. Alexander S. Behr * a, Diana Chernenko a, Dominik Koßmann b, Arjun Neyyathala c, Schirin Hanf c, Stephan A. Schunk d and Norbert Kockmann a a Department of Biochemical and Chemical Engineering, Laboratory of Equipment Design, TU Dortmund University, Dortmund, Germany.
Male sex, early life chemical exposure and the brain aromatase enzyme have been implicated in autism spectrum disorder (ASD). In the Barwon Infant Study birth cohort (n = 1074), higher prenatal ...
Increased consumption of ultra-processed foods (UPFs) which have additives such as artificial colours, flavours and are usually high in salt, sugar, fats and specific preservatives, are associated with diet-related non-communicable diseases (NCDs). In India, there are no standard criteria for identifying UPFs using a classification system based on extent and purpose of industrial processing.